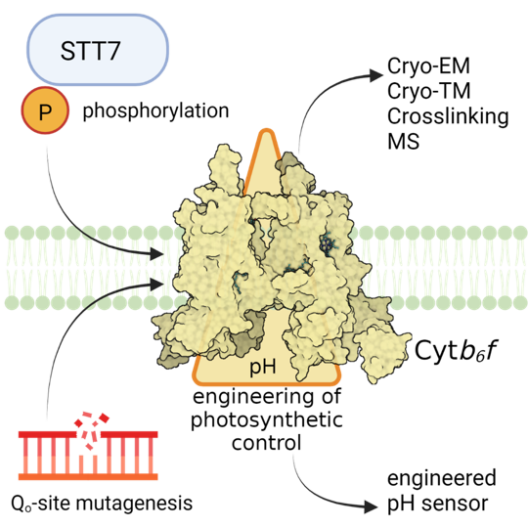
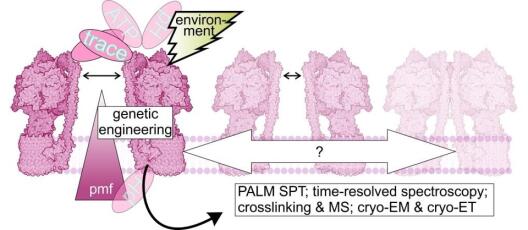
In photosynthetic electron transfer, the cytochrome b6f complex (Cyt b6f) contributes to the generation of both PMF components, ΔpH and ΔΨ. This process is driven by the Q cycle and operates in two distinct modes – via the canonical Q cycle during linear electron flow (LEF) and via an alternative Q cycle during cyclic electron flow (CEF), attributing a ferredoxin-plastoquinone reductase activity to the Cyt b6f. Project 1 aims to provide mechanistic insights into the structure-function tuning of the lumenal Qo-site and the stromal Qi-site of Cyt b6f. The Qo-site is responsible for PQH2 oxidation and electron bifurcation as well as for pH-dependent slowdown of photosynthetic electron transfer. This regulatory process is also known as “photosynthetic control”, based on a still unknown molecular mechanism. The Qi-site in Cyt b6f is important for acceptance of stromal electrons during CEF, concomitant with switching of the Q cycle from the canonical to the alternative mode. Utilizing the green algal model species Chlamydomonas reinhardtii, Project 1 endeavors to determine the role of STT7, a thylakoid-associated Ser/Thr protein kinase, and/or phosphorylation(s) of Cyt b6f subunit-IV and subunit PETO for structure-function tuning. Furthermore, Project 1 aims to reveal molecular mechanisms contributing to photosynthetic control at the Qo site through site directed mutagenesis of candidate residues. Moreover, Project 1 plans to perform comparative cryo-EM structural analyses, including chemical crosslinking and mass spectrometry, of isolated wild type and mutant Cyt b6f. Project 1 further aims to structurally elucidate antimycin-A (AA) binding to Cyt b6f and to investigate native thylakoid membranes of wild type and mutant Cyt b6f in the presence and absence of AA via cryo-ET (together with Project 8). Moreover, Project 1 intends to engineer Cyt b6f as a sensor of chloroplast PMF (together with the Central Project).
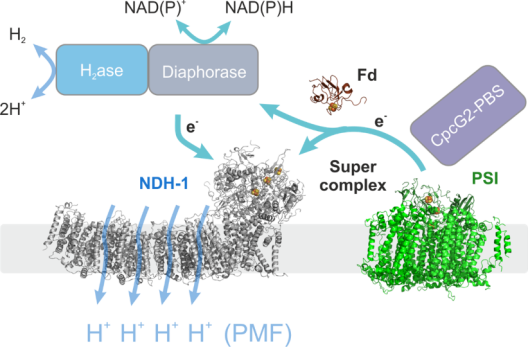
Membrane protein complexes play a crucial role in bioenergetics, particularly in photosynthetic organisms. They are responsible for the light-to-chemical energy conversion in the primary reactions of photosynthesis, thereby driving the energy gradient that promotes and sustains almost all life on our planet. In this fundamental process, they combine the light driven transport of electrons with the vectorial transport of protons across a membrane. Project 2 focuses on one of these molecular machines, complex I (NDH-1), which is part of photosynthetic as well as respiratory electron transport chains. All NDH-1 versions combine electron transfer from ferredoxin (Fd) to plastoquinone with the transfer of protons across the thylakoid membrane in a yet unknown fashion. Many questions concerning their precise molecular function remain unanswered due to the lack of structure-function based mutagenesis studies in vitro and in vivo. In principle, as highly efficient proton pumps, they could contribute to light-driven proton motive force (PMF) formation substantially by accepting electrons from photosystem I (PSI) via Fd in cyclic electron transport, but their role in vivo is rather unclear. Project 2 provides novel molecular insights into NDH-1 function and its integration into the dynamic electron transport network of the cell based on recent advances in structural analyses of the cyanobacterial complex. Specifically, Project 2 focuses on supercomplex formation with PSI along with CpcG2-phycobilisomes and with H2ase/Diaphorase (together with Project 3) to understand its role in PMF formation. Project 2 aims to investigate cyanobacterial supercomplexes in the model organisms Synechocystis sp. PCC 6803 and the filamentous, diazotrophic cyanobacterium Anabaena sp. PCC 7120, which will be compared to the land plant supercomplex form the bryophyte Physcomitrella patens, a genetically accessible system that allows comparatively easy upscaling of biomass production.
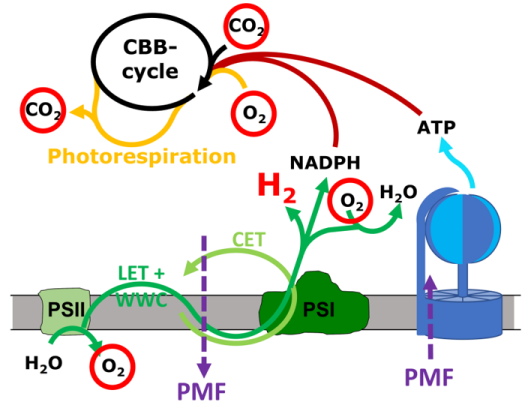
Cyanobacteria harbor a diversity of membrane protein complexes in their thylakoid membrane that are involved in respiratory electron transport (RET), linear and cyclic electron transport (LET and CET) and the water-water cycle (WWC). These intertwined routes offer a great flexibility to the system that enables the cells to rapidly respond to changing light intensities and other environmental conditions. Although many of the involved complexes are known in detail, the relative contribution of the different routes, i.e. the way the cells convert the absorbed light energy into a usable output like the proton motive force (PMF, in turn ATP) and NADPH, is unclear. Project 3 aims to quantify the rates of electron flow through PSII and PSI simultaneously with O2 evolution and consumption as well as CO2 fixation and liberation processes in the cyanobacterium Synechocystis sp. PCC 6803. This is achieved by coupling a mass spectrometer to the recently developed Dual-KLAS-NIR instrument. Adding isotopically labeled O2 and CO2 it is possible to discern the CBB cycle, photorespiration and O2 evolution by PSII from its consumption by the WWC. The collected data will deliver a complete picture of the involved electron transport routes, the amount of ATP and NADPH produced and consequently also a measure of the PMF in play. Project 3 also plans to complement these measurements by expressing pH sensitive fluorescent proteins in the cyanobacterial cytosol and thylakoid lumen (together with Project 7). Knowledge on PMF formation and its potential dissipation in the cells is essential for any efficient future use of the photosynthetic electron transport chain in terms of biological H2 production. If a large proportion of the electrons derived from water oxidation is diverted to H2 production, an imbalance of PMF formation compared to NADPH production is predetermined. To maximize photosynthetic H2 production and to develop a persisting process, it is imperative to dissipate part of the PMF. Project 3 works with a recently discovered mutant strain that diverts part of its PMF to heat, aiming to investigate the mechanisms and proteins involved as a basis for future applications in strains optimized for light-driven H2 production. Cyanobacterial bidirectional hydrogenase receives electrons from a variety of different substrates, including H2, NADH, NADPH, ferredoxin and flavodoxin. Project 3 investigates in vitro whether electron delivery from hydrogenase to photosynthetic complex I (NDH-1) occurs via direct binding of hydrogenase to NDH-1 (together with Project 2) or is mediated by an electron carrier.
The thylakoid ultrastructure of the green alga Chlamydomonas reinhardtii provides dynamic sub-compartments where specific alterations in ATP synthase activity and its spatiotemporal organization could occur. ATP synthase is a major bioenergetic complex that affects H+ usage, changes the PMF and eventually impacts photosynthetic electron transfer regulation. The objective of Project 4 is to dissect the tight relation between photosynthesis and ATP synthase activity along with the effect of spatiotemporal features, in particular the dynamic ultrastructure of thylakoids and the localization and mobility of ATP synthase within. Together with the Central Project, Project 4 engineers ATP synthase at different sites with respective sensors for ATP and pH. Addition of fluorescent sensors or tags for single molecule labeling facilitates tracing of individual enzymes and measuring of local bioenergetics parameters. Project 4 aims to monitor tagged ATP synthase under various cellular conditions and to establish a potential correlation between its recorded activity and possible changes in its molecular organization and/or sub-compartmental location. Project 4 determines enzyme activity by established time-resolved optical methods and through novel high-resolution ratiometric readouts of fluorescent sensors for pH and ATP. Moreover, Project 4 records ATP synthase spatiotemporal organization and changes in protein environment by fluorescence lifetime imaging and single particle tracking in combination with biochemical analyses. Project 4 investigates impacts on the thylakoid ultrastructure along with details of the protein complex itself and yet unknown interactors via cryogenic electron microscopy (together with Project 8) and mass spectrometry as well as analyses their relevance for the PMF amplitude in light and darkness. A sensor equipped ATP synthase represents a valuable tool, allowing for microscopic quantifications of both ATP synthase substrates – the electrochemical PMF (as ΔpH) and the chemical adenylate status of the cell (as ATP).
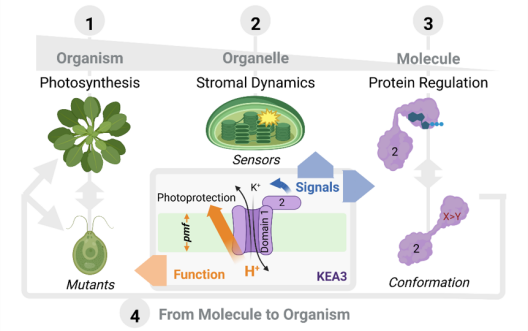
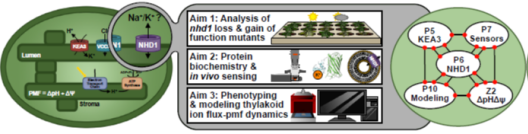
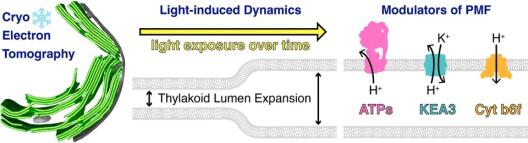
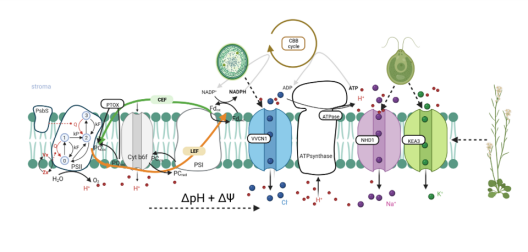
The chloroplast PMF is produced via light-driven proton-coupled electron transport across the thylakoid membrane. This generates both a membrane potential (Δψ) and a proton concentration gradient (ΔpH). Their contribution to the PMF is governed in part by thylakoid ion transport processes. The lumenal proton concentration determines the fraction of light energy that is used for photochemistry. The thylakoid K+-exchange antiporter 3 (KEA3) plays a fundamental role for the flexibility of vascular plant photosynthesis during light fluctuations. KEA3 senses the energy state of the chloroplast stroma and adjusts the levels of the photoprotective energy dependent quenching (qE) in response. If the energy state is low, KEA3 is activated to decrease the lumenal proton concentration shut off qE. While KEA3 activity affects the PMF composition and decreases the ΔpH component, it appears to have little effect on the PMF size and thus its energetic function. Deregulation of KEA3 activity can result in transiently higher CO2 fixation rates. This proposes KEA3 regulation as a putative target to enhance photosynthesis under the dynamic light conditions found in the field. An in-depth understanding of KEA3 regulation and its interaction with the complex network of photosynthetic reactions will be pivotal to further explore this avenue. Project 5 explores KEA3 function in PMF partitioning and its regulation via a comparative approach: By measuring KEA3 activity in the unicellular alga Chlamydomonas reinhardtii, Project 5 aims to understand the evolution of KEA3 function and regulation. Moreover, Project 5 characterizes how environmental changes affect the chloroplast energy state by comparing stromal energy dynamics and their relation to KEA3 activity between Arabidopsis thaliana and C. reinhardtii. By expressing modified KEA3 versions in planta, Project 5 intends to confirm KEA3 regulating factors and attempts to improve photosynthetic performance and growth.
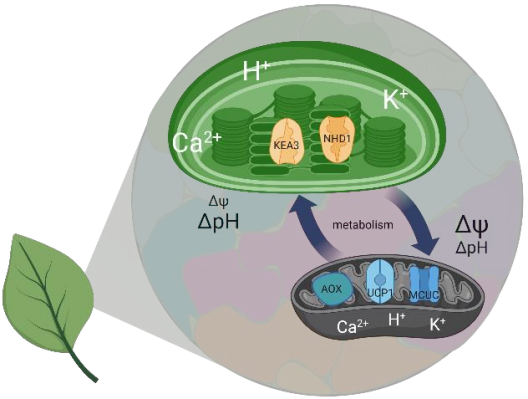
Project 6: Studying the functional significance of the putative Na+/H+ carrier AtNHD1 in the thylakoid membrane for proton motive force dynamic and partitioning
The PMF is partitioned into two energetically equivalent components, a pH gradient (ΔpH) and a membrane potential (ΔΨ). Their relative contribution is dependent on the light intensity and its dynamics. Low lumenal pH has important protective implications, decreasing e- transport via the cytochrome b6f complex and activating energy dissipation as heat (qE), the main component of non-photochemical quenching (NPQ). Plants strike a balance between photoprotection and biomass gain by finetuning PMF partitioning. Ion transport proteins in the thylakoid membrane play a key role in this regard because H+ transfer into the lumen is coupled to a counter-ion flux that dissipates the membrane potential to allow for lumenal H+ accumulation. It follows that unraveling the thylakoid ion transportome is a prerequisite for our understanding of the PMF and of how to improve photosynthetic productivity. In the model plant Arabidopsis thaliana, transport proteins mediating Cl- flux (VCCN1) and K+/H+ exchange (KEA3) participate in PMF partitioning. At least one additional mediator for K+ or monovalent cations is expected in the thylakoid membrane. Project 6 focuses on NHD1, a putative Na+/H+ exchanger residing in the thylakoid membrane. Heterologous expression of NHD1 restores growth of E. coli mutants defective in K+ transport, indicating NHD1 might facilitate K+ flux across thylakoid membranes in vivo. Project 6 seeks to determine the consequences caused by loss and gain of NHD1 function on PMF, photosynthesis, and global gene expression using Arabidopsis thaliana; to characterize NHD1 substrate specificity; and to design and spectroscopically analyse higher order mutants devoid of NHD1, VCCN1, and KEA3 to update the current model of PMF dynamics and its molecular players under fluctuating light conditions.
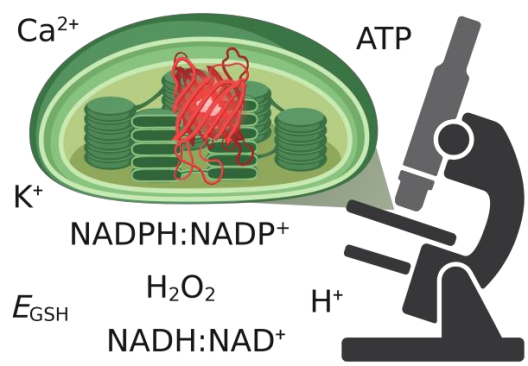
Mitochondria and chloroplasts have been shaping eukaryotic life as we know it. At the center of the function of both organelles is the proton motive force (PMF). The PMF connects redox chemistry of electron transport with ATP synthesis, but also acts as a regulation hub that determines the function and the makeup of both organelles. In plants and algae, the PMF of both mitochondria and chloroplasts are interconnected via the cellular metabolic network. While efficient photosynthesis requires chloroplast and mitochondrial bioenergetics to work in concert, how exactly this interplay is achieved in vivo and which proteins contribute to which extent at the level of the PMF remains insufficiently understood. Project 7 elucidates how the subcellular dynamics of protons (H+), potassium ions (K+) and calcium ions (Ca2+) determine PMF characteristics in mitochondria and chloroplasts in vivo. Using a mature rosette leaf of Arabidopsis thaliana as a model, Project 7 exploits a set of fluorescent protein-based biosensors and in vivo high-resolution imaging to generate an atlas of H+, K+ and Ca2+ dynamics in the subcompartments of chloroplasts and mitochondria in folio. This atlas will not only comprise three-dimensional molecular information in living leaf tissues, but also responses to different illumination regimes. It will provide a hitherto unique reference and will be used to further corroborate the significance of candidate proteins for H+, K+ and Ca2+ transport across the organelle membranes in determining PMF characteristics in folio. Project 7 tests a selection of genetic mutants with functional defects of mitochondrial and chloroplast H+, K+ and Ca2+ dynamics for their impact on organellar PMF characteristics in folio. These candidate proteins include KEA3 (together with Project 5) and NHD1 (together with Project 6) as well as the MCUs, MICU, UCP1 and AOX1a. Combining the atlas approach with the mutant analysis allows to address the central question to what extent changes in PMF characteristics in one organelle affect the other. The significance of altered PMF dynamics for photosynthetic metabolism, organelle ultrastructure, and whole plant performance will become accessible. An advanced understanding of PMF regulation in vivo and the relative significance of specific candidate protein players in setting PMF characteristics will be established as a result.
Thylakoid membranes scaffold photosynthetic complexes, which couple electron transfer to proton movement across the membrane, thereby generating the proton motive force (PMF, composed of ΔpH and ΔΨ) that drives the production of ATP. Thylakoid architecture is intimately linked to efficient photosynthesis. Thylakoids are flat instead of spherical, increasing the concentration of photosynthetic complexes per lumenal area to drive the PMF, while providing directionality for the diffusion of the soluble electron carrier plastocyanin through the thylakoid lumen. However, thylakoid architecture is not static and responds to changes in light quality and quantity. Preliminary electron microscopy observations in cyanobacteria, green algae, and plants have shown that the thylakoid lumen swells under increased light intensity. A logical hypothesis would be that lumen acidification (ΔpH) and ion flux-driven changes in membrane potential (ΔΨ) may be coupled to osmotic forces that expand the lumen. However, the precise dynamics and mechanistic details of this architectural change remain uncharacterized. The goal of Project 8 is to understand the mechanistic basis and physiological consequences of light-induced thylakoid swelling, and to specifically test how this swelling is modulated by different components of the PMF. This is enabled by an advanced cryo-electron tomography (cryo-ET) workflow, developed to measure native thylakoid architecture (membrane and lumen width) with sub-nanometer precision and to analyze the organization of each photosynthetic complex along these membranes. First, project 8 aims to characterize the kinetics of thylakoid swelling in green algae (Chlamydomonas reinhardtii) and cyanobacteria (Synechocystis sp. 6803) by performing a detailed cryo-ET time-series of wild type cells after transition from darkness to different light intensities. Next, Project 8 intends to examine the contributions of ΔpH and ΔΨ to thylakoid lumen expansion by combining cryo-ET with small molecule ionophores and genetic perturbations of ATP synthase (together with Project 4) and KEA3 ion antiporters (together with Project 5). In parallel, Project 8 endeavors to test the specific PMF contributions of cytochrome b6f to thylakoid lumen dynamics (together with Project 1). Finally, Project 8 aims to create an integrated model of light-dependent lumen expansion by combining cryo-ET data with functional measurements of PMF and photosynthetic function.
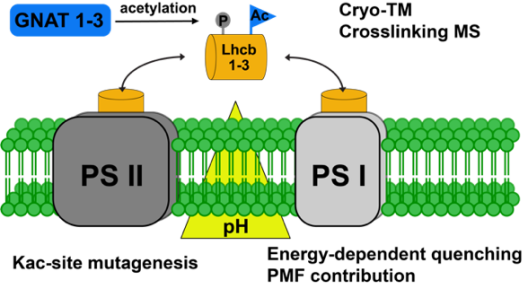
Photosynthetic electron transport is dynamically regulated in response to environmental conditions to prevent photoinhibition and photodamage of the photosystems. Several chemical modifications of proteins are known to play important roles in modulating photosynthetic yield during plant acclimation responses. Next to phosphorylation, acetylation of amino groups within proteins is a major posttranslational modification in all organisms and is particularly abundant on proteins involved in photosynthesis. N-terminal acetyltransferases or lysine acetyltransferases catalyze the transfer of the acetyl group from acetyl coenzyme A to the targeted amino group. In contrast to N-terminal acetylation, the acetylation of lysine residues (KAc) is reversible, whereby lysine deacetylases are responsible for the removal of acetyl groups from proteins. Recently, eight new dual-specific acetyltransferases (GNATs) were identified in chloroplasts of A. thaliana. The first characterization of GNAT2 revealed that this enzyme is indispensable for photosynthetic state transitions. In addition, the mutants show increased energy-dependent quenching at high light intensities compared to wild type. While the first protein substrates of GNAT2 have been identified, the specific role of protein acetylation in the acclimation responses of the photosynthetic light reactions is still not understood on a mechanistic level. Project 9 aims to provide novel biochemical insights into the functions of acetylation on selected target proteins, as well as the involvement of the closely related plastidial acetyltransferases GNAT1-3 in regulating acetylation of photosynthetic proteins contributing to energy-dependent quenching. Project 9 implements latest in-gel chemical crosslinking techniques in combination with cryo-electron tomography (together with Project 8) for 3D modelling to study differences in thylakoid protein complex composition dependent on light and genotype. Specifically, Project 9 focuses on the role of acetylation in the photosynthetic acclimation responses to changes in light availability.
The proton motive force (PMF) across the thylakoid membrane plays a pivotal role in the regulation of photosynthetic organisms’ energy metabolism. The number of interdependent processes and possible cross-talks between reactions that generate, modulate and depend on the PMF are astonishing and their combined effect on the cell’s bioenergetics leads to a nonlinear complexity. Hence, a rigorous framework that allows for a systematic disentangling of this complexity is necessary to develop concepts on how the PMF is achieved and regulated across photosynthetic species. Computational, mechanistic models of the photosynthetic electron transport chain offer a quantitative framework that enables comparative studies. Numerous models of photosynthesis describe the PMF, but many of them focus solely on its pH-dependent component (ΔpH), mainly due to its link to the important photoprotective mechanism named non-photochemical quenching. Energy-dependent quenching releases excess light energy as heat, to protect photosystem II against damage. Yet, the importance of the membrane potential (ΔΨ) in the generation of the PMF and photoprotection should not be ignored, as photosynthetic electron transport is highly sensitive to the concentration of ions (e.g., H+, K+, Mg2+ and Cl−). Ion transport proteins in the thylakoid membrane, such as KEA3, allow for an out-flux of H+ with a counter-in-flux of cations, affecting the contribution of proton-based or potential-driven components of the PMF. By dynamic fine-tuning of the two PMF components, photosynthetic organisms can dynamically adapt to environmental changes and balance their energy production to meet their energy demand. Project 10 aims to construct a unifying theoretical framework describing the dynamics of the PMF for numerous classes of photosynthetic organisms studied within GoPMF, ranging from cyanobacteria to green microalgae to vascular plants. Building on previously constructed mathematical models of the photosynthetic electron transport chain, Project 10 fully explores the modular design of these models to create a blueprint for PMF regulation. Within this framework, Project 10 systematically investigates the effect of PMF partitioning on photosynthetic dynamics and performs cross-species comparisons to generate novel hypotheses regarding PMF regulation. Furthermore, combined with the collected experimental work of GoPMF, the model will guide experiments investigating the light-driven production of hydrogen in cyanobacteria.
Rapid advances in high-resolution imaging and the expansion of in vivo biosensing have been revolutionizing biological research. The power of both approaches combined allows for the quantification of cellular physiology in situ and the visualization of (sub-)organellar and molecular dynamics. Those developments have recently also started to change our understanding of the proton motive force (PMF). Visualization of pH dynamics and Δψ in mitochondria has painted an unexpected picture of how the PMF is organized and how it changes dynamically in single organelles. Comparable advances for the PMF across photosynthetic membranes are yet to be made and dedicated method development is required. Understanding PMF dynamics in vivo further demands capabilities for live sensing of ions, such as K+, that contribute to PMF partitioning, and associated metabolites, such as NADPH:NADP+, NADH:NAD+ and ATP, that are tightly linked with the PMF of chloroplasts. Since many of the now available biosensors are protein-based, specific subcellular targeting and genetic fusion to proteins in bioenergetic membranes is feasible to measure with highest local specificity. However, implementation of these biosensors requires extensive characterization of their particular biochemical and biophysical properties in isolation and in situ to ensure specificity and to avoid artefacts. Specialized technical knowledge is a prerequisite that is unrealistic to be acquired by each individual user. The Central Project brings together the available expertise on biosensing with new experimental findings. Furthermore, the Central Project develops biosensing strategies for photosynthetic PMF monitoring and optimizes high resolution light microscopy of chloroplasts as well as photosynthetic membranes. Specifically, the Central Project establishes stromal and lumenal biosensing of H+ to obtain a direct readout of ΔpH. Moreover, the Central Project pioneers future projects with NADPH:NADP+ biosensing and optimizes bespoke setups for biosensing under varying photosynthetic activities. Furthermore, the Central Project aims to develop super-resolution imaging with sub-organellar resolution for photosynthetic membranes. This is not limited to live imaging by confocal microscopy with state-of-the-art deconvolution and Airyscan detection, but also includes advanced protocols of protein labeling for super-resolution imaging, single particle localization microscopy and mobility studies of thylakoid membrane proteins. In close collaboration with members and projects of GoPMF, the Central Project continuously extends, refines and implements the toolbox for the optical analysis of PMF determinants.