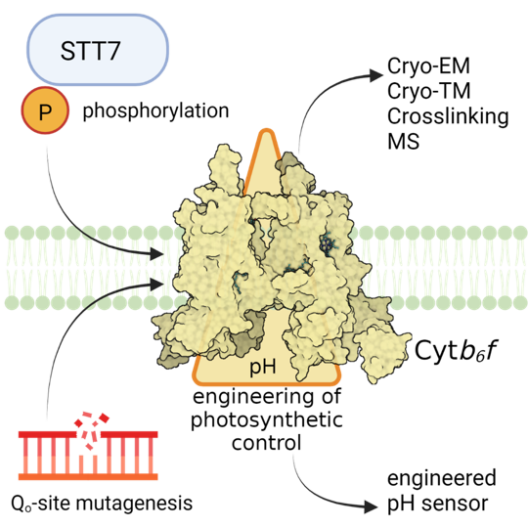
Im Rahmen des photosynthetischen Elektronentransfers trägt der Cytochrom b6f Komplex (Cyt b6f) zur Ausbildung beider PMF-Komponenten bei, ΔpH und ΔΨ. Dieser Prozess wird vom Q-Zyklus angetrieben und läuft in zwei unterschiedlichen Modi ab – über den kanonischen Q-Zyklus während des linearen Elektronentransfers (LEF) und über einen alternativen Q-Zyklus während des zyklischen Elektronentransfers (CEF), der dem Cyt b6f eine Ferredoxin-Plastochinon-Reduktase-Aktivität zuschreibt. Projekt 1 hat zum Ziel, mechanistische Einblicke in die Struktur-Funktions-Abstimmung der lumenalen Qo-Stelle und der stromalen Qi-Stelle des Cyt b6f zu liefern. Die Qo-Stelle ist für die PQH2-Oxidation und die Elektronenbifurkation sowie für die pH-abhängige Verlangsamung des photosynthetischen Elektronentransfers verantwortlich. Dieser regulatorische Prozess ist auch als „photosynthetische Kontrolle“ bekannt, basierend auf einem im Detail noch unbekannten molekularen Mechanismus. Die Qi-Stelle im Cyt b6f ist für die Aufnahme von stromalen Elektronen während des CEF wichtig, einhergehend mit dem Umschalten des Q-Zyklus vom kanonischen zum alternativen Modus. Mit Hilfe des Grünalgen Modellorganismus Chlamydomonas reinhardtii hat Projekt 1 zum Ziel, die Rolle der Thylakoid-assoziierten Ser/Thr-Proteinkinase STT7 und/oder von Phosphorylierung(en) der Cyt b6f Untereinheit IV sowie der Untereinheit PETO für die Struktur-Funktions-Abstimmung zu untersuchen. Außerdem beabsichtigt Projekt 1 mittels ortsgerichteter Mutagenese von Kandidatenresten molekulare Mechanismen zu identifizieren, die zur photosynthetischen Kontrolle an der Qo-Stelle beitragen. Darüber hinaus plant Projekt 1 vergleichende Kryo-EM-Strukturanalysen, einschließlich chemischer Vernetzung und massenspektrometrischer Analysen, mit isoliertem Wildtyp- und Mutanten-Cyt b6f durchzuführen. Ferner hat Projekt 1 zum Ziel, die Antimycin-A (AA) Binding im Cyt b6f strukturell aufzuklären und native Thylakoidmembranen von Wildtyp- und Mutanten-Cyt b6f in An- und Abwesenheit von AA mittels Kryo-ET zu untersuchen (gemeinsam mit Projekt 8). Zusätzlich beabsichtigt Projekt 1 Cyt b6f als Sensor für die chloroplastidäre PMF zu konstruieren (gemeinsam mit dem Zentralprojekt).
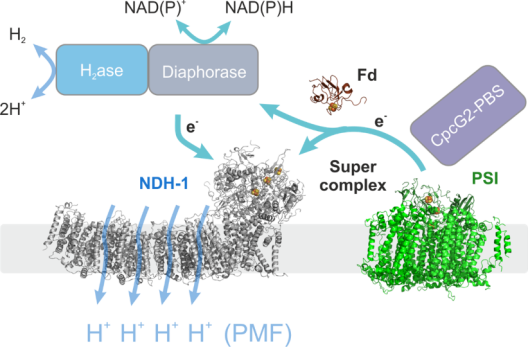
Membranproteinkomplexe spielen eine entscheidende Rolle in der Bioenergetik, insbesondere in photosynthetischen Organismen. Sie sind für die Umwandlung von Licht in chemische Energie in der Primärreaktion der Photosynthese verantwortlich und treiben so den Energiegradienten an, der für fast alles Leben auf unserem Planeten verantwortlich ist. In diesem grundlegenden Prozess kombinieren sie den lichtgetriebenen Transport von Elektronen mit dem vektoriellen Transport von Protonen über eine Membran. Projekt 2 ist auf einen Typ dieser molekularen Maschinen fokussiert, den Komplex I (auch bekannt als NDH-1), der sowohl Teil der photosynthetischen als auch der respiratorischen Elektronentransportkette ist. Alle NDH-1 Varianten verbinden den Elektronentransfer von Ferredoxin (Fd) zu Plastochinon mit dem Transfer von Protonen durch die Thylakoidmembran auf bisher unbekannte Weise. Viele Fragen zur genauen molekularen Funktion sind noch unbeantwortet, da es keine auf Struktur-Funktions-Mutagenese basierenden in vitro und in vivo Studien gibt. Als hocheffiziente Protonenpumpe könnte der NDH-1 Komplex wesentlich zur Bildung der protonenmotorischen Kraft (PMF) beitragen, indem er im zyklischen Elektronentransport Elektronen von Photosystem I (PSI) über Fd aufnimmt, aber seine Rolle in vivo ist eher unklar. Projekt 2 liefert neue molekulare Einblicke in die Funktion von NDH-1 und seine Einbindung in das dynamische Elektronentransportnetzwerk der Zelle, basierend auf aktuellen Fortschritten in der Strukturanalyse des cyanobakteriellen Komplexes. Insbesondere ist Projekt 2 auf die Superkomplexbildung mit PSI und CpCG2-Phycobilisomen sowie mit H2ase/Diaphorase (gemeinsam mit Projekt 3) fokussiert, um seine Rolle bei der PMF-Bildung zu verstehen. Projekt 2 hat zum Ziel cyanobakterielle Superkomplexe in den Modellorganismen Synechocystis sp. PCC 6803 und dem filamentösen, diazotrophen Cyanobakterium Anabaena sp. PCC 7120 zu untersuchen, und diese mit dem Landpflanzen-Superkomplex des Bryophyten Physcomitrella patens zu vergleichen, einem genetisch zugänglichen System, das eine vergleichsweise einfache Hochskalierung der Biomasseproduktion ermöglicht.
Projekt 3: Untersuchungen zur protonenmotorischen Kraft (PMF) im Cyanobakterium Synechocystis mit dem Ziel die photosynthetische Wasserstoffproduktion zu maximieren
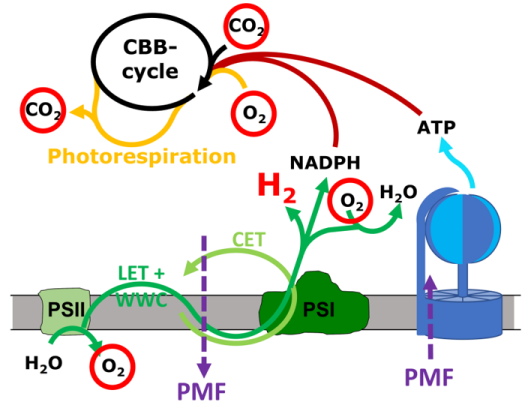
Cyanobakterien beherbergen eine Vielfalt von Membranproteinkomplexen in ihrer Thylakoidmembran, die am respiratorischen Elektronentransport (RET), am linearen und zyklischen Elektronentransport (LET und CET) und am Wasser-Wasser-Zyklus (WWC) beteiligt sind. Diese miteinander verflochtenen Wege bieten dem System eine große Flexibilität, die es den Zellen ermöglicht, schnell auf sich ändernde Lichtintensitäten und andere Umweltbedingungen zu reagieren. Obwohl viele der beteiligten Komplexe im Detail bekannt sind, ist der relative Beitrag der verschiedenen Wege, d.h. die Art und Weise, wie die Zellen die absorbierte Lichtenergie in eine nutzbare Form wie die protonenmotorische Kraft (PMF, und damit ATP) und NADPH umwandeln, unklar. Projekt 3 hat zum Ziel die Elektronentransferraten durch Photosystem II und I (PSII und PSI) parallel zu O2 Entwicklung und Verbrauch sowie und CO2 Fixierung und Freisetzung im Cyanobakterium Synechocystis sp. PCC 6803 zu quantifizieren. Dies wird durch die Kopplung eines Massenspektrometers an das kürzlich entwickelte Dual-KLAS-NIR-Instrument erreicht. Durch die Zugabe von isotopenmarkiertem O2 und CO2 ist es möglich, den CBB-Zyklus, die Photorespiration und die O2-Evolution durch PSII von dessen Verbrauch durch den WWC zu unterscheiden. Die gesammelten Daten werden ein vollständiges Bild über die involvierten photosynthetischen Elektronentransferrouten sowie die Mengen an produziertem ATP und NADPH liefern. Daraus können Aussagen zum Protonengradienten und zur protonenmotorischen Kraft (PMF) abgeleitet werden. Projekt 3 plant auch diese Messungen durch die Expression pH-sensitiver fluoreszierender Proteine im cyanobakteriellen Zytosol und Thylakoidlumen zu ergänzen (gemeinsam mit Projekt 7). Wissen zur Erzeugung und möglichen Dissipation der PMF ist essenziell, um die photosynthetische Elektronentransportkette zukünftig effizient für biologische Wasserstoffproduktion nutzen zu können. Wenn ein großer Anteil der Elektronen aus der Wasserspaltung für die Synthese von H2 genutzt wird, führt dies zu einem Ungleichgewicht zwischen PMF-Bildung (ATP-Synthese) und NADPH-Produktion. Um die photosynthetische Wasserstoffproduktion zu maximieren und einen anhaltenden Prozess zu entwickeln, ist es daher notwending die PMF teilweise abzubauen. Projekt 3 arbeitet mit einem kürzlich entdeckten Mutantenstamm, der einen Teil seiner protonenmotorischen Kraft (PMF) als Wärme ableitet. Dies hat zum Ziel, die an diesem Vorgang beteiligten Mechanismen und Proteine zu untersuchen, als Grundlage für zukünftige Anwendungen in Stämmen, die für die Licht-getriebene H2-Produktion optimiert sind. Die bidirektionale Hydrogenase der Cyanobakterien erhält Elektronen von einer Vielzahl verschiedener Substrate, darunter H2, NADH, NADPH, Ferredoxin und Flavodoxin. Projekt 3 untersucht in vitro, ob die Elektronenübertragung von der Hydrogenase zum photosynthetischen Komplex I (NDH-1) direkt durch Bindung der Hydrogenase an NDH-1 erfolgt (gemeinsam mit Projekt 2) oder ob sie durch einen Elektronenüberträger vermittelt wird.
Die Ultrastruktur der Thylakoidmembran in der Grünalge Chlamydomonas reinhardtii beinhaltet Unterkompartimente, in denen spezifische Veränderungen der ATP-Synthaseaktivität und ihrer räumlich-zeitlichen Organisation auftreten können. Die ATP-Synthase ist ein wichtiger bioenergetischer Komplex, der den H+-Fluss durch die Membran treibt, die protonenmotorische Kraft (PMF) verändert und schließlich die Regulierung des photosynthetischen Elektronentransfers beeinflusst. Projekt 4 hat zum Ziel, zum einen die enge Beziehung zwischen photosynthetischer Effizienz und ATP-Synthaseaktivität zu beleuchten, und zum anderen die gleichzeitigen Auswirkungen raum-zeitlicher Merkmale zu untersuchen. Die dynamische Ultrastruktur der Thylakoide sowie die Lokalisierung und Mobilität der ATP-Synthase nimmt dabei einen besonderen Stellenwert ein. Zu diesem Zweck wird die ATP-Synthase in Zusammenarbeit mit dem Zentralprojekt an verschiedenen Stellen mit entsprechenden Sensoren für ATP und pH ausgestattet. Das Anbringen von Fluoreszenzsensoren oder Molekülmarkierungen ermöglicht die Verfolgung einzelner Enzyme und die Messung lokaler bioenergetischer Parameter. Projekt 4 zielt darauf ab, markierte ATP-Synthase unter verschiedenen zellulären Bedingungen zu überwachen und eine mögliche Korrelation zwischen ihrer aufgezeichneten Aktivität und potentiellen Veränderungen ihrer molekularen Organisation und/oder ihrer subkompartimentellen Lage herzustellen. Projekt 4 bestimmt die Enzymaktivität mit Hilfe etablierter zeitaufgelöster optischer Methoden und neuartiger hochauflösender ratiometrischer Erfassung von Fluoreszenzsensoren für pH und ATP bestimmt. Darüber hinaus untersucht Projekt 4 die räumlich-zeitliche Organisation und Veränderungen in der Proteinumgebung durch Halbwertszeitmessungen der Fluoreszenz und Einzelpartikelverfolgung in Kombination mit biochemischen Analysen. Projekt 4 erfasst Auswirkungen auf die Thylakoid-Ultrastruktur und Details des Proteinkomplexes selbst sowie bisher unbekannter Interaktoren mittels kryogener Elektronenmikroskopie (gemeinsam mit Projekt 8) oder Massenspektrometrie und testet ihre Relevanz für die PMF-Amplitude in Licht und Dunkelheit. Eine mit einem Sensor ausgestattete ATP-Synthase stellt ein wertvolles Werkzeug dar, das die mikroskopische Quantifizierung beider ATP-Synthasesubstrate - der elektrochemischen PMF (als pH-Wert) und des chemischen Adenylat-Status der Zelle (als ATP) - ermöglichen wird.
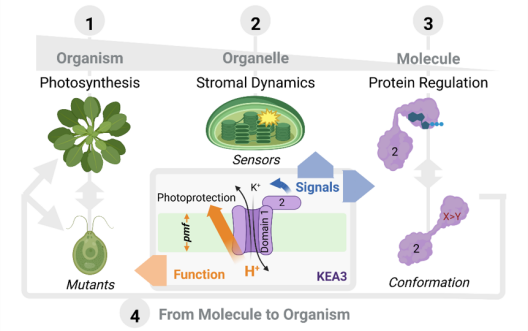
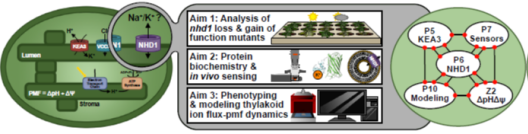
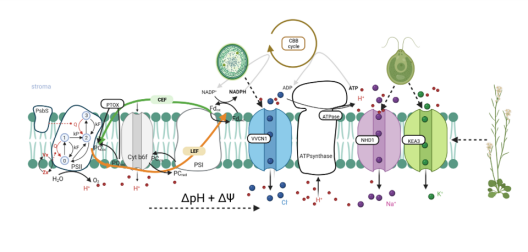
Die chloroplastidäre PMF wird durch lichtgetriebenen, protonengekoppelten Elektronentransport über die Thylakoidmembran erzeugt. Dies produziert sowohl ein Membranpotential (∆ψ) als auch einen Protonenkonzentrationsgradienten (∆pH), deren Beiträge zur PMF zum Teil von Thylakoid-Ionentransportprozessen bestimmt werden. Die lumenale Protonenkonzentration bestimmt den Anteil der Lichtenergie, welcher für Photochemie genutzt wird. Der K+-Austausch-Antiporter 3 (KEA3) in der Thylakoidmembran spielt eine grundlegende Rolle für die Flexibilität der Photosynthese von Gefäßpflanzen bei Lichtschwankungen. KEA3 erkennt den Energiezustand des Chloroplastenstromas und passt als Reaktion darauf das Ausmaß des photoprotektiven energieabhängigen Quenchings (qE) an. Wenn der Energiezustand niedrig ist, wird KEA3 aktiviert um die Protonenkonzentration im Lumen zu verringern und somit qE abzuschalten. Während KEA3 die PMF-Zusammensetzung beeinflusst und die ∆pH-Komponente verringert, scheint es nur geringe Auswirkungen auf die PMF-Größe und damit ihre energetische Funktion zu haben. Eine De-Regulierung der KEA3-Aktivität kann zu vorübergehend höheren CO2-Fixierungsraten führen. Dies deutet darauf hin, dass KEA3-Regulierung ein mögliches Ziel für die Verbesserung der Photosynthese unter den im Freiland herrschenden dynamischen Lichtbedingungen darstellt. Ein tiefgreifendes Verständnis der KEA3-Regulierung und ihrer Interaktion mit dem komplexen Netzwerk photosynthetischer Reaktionen wird für die weitere Verfolgung dieses Ansatzes von entscheidender Bedeutung sein. Projekt 5 erforscht die Funktion von KEA3 bei der PMF-Verteilung und deren Regulierung durch einen vergleichenden Ansatz: Durch Messungen der KEA3-Aktivität in der einzelligen Alge Chlamydomonas reinhardtii beabsichtigt Projekt 5 evolutionäre Auswirkungen auf die KEA3-Funktion und -Regulierung zu verstehen. Darüber hinaus charakterisiert Projekt 5 durch den Vergleich stromaler Energiedynamiken und ihrer Verbindungen mit der KEA3-Aktivität zwischen Arabidopsis thaliana und C. reinhardtii, wie sich Umweltveränderungen auf den chloroplastidären Energiezustand auswirken. Durch die Expression modifizierter KEA3-Versionen in planta hat Projekt 5 außerdem zum Ziel, KEA3-regulierende Faktoren zu bestätigen und zu versuchen, photosynthetische Leistung und Pflanzenwachstum zu verbessern.
Projekt 6: Untersuchungen zur Wichtigkeit des putativen Na+/H+ Austauschers AtNHD1 für Aufbau und Komposition der plastidären protonenmotorischen Kraft
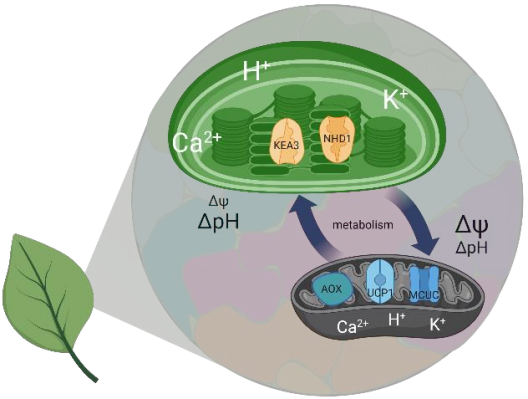
Die PMF ist in zwei energetisch äquivalente Komponenten geteilt, einen pH-Gradienten (ΔpH) und ein Membranpotential (Δψ). Ihr relativer Beitrag ist abhängig von der Lichtintensität und ihrer Dynamik. Ein niedriger pH-Wert hat auch eine wichtige Schutzfunktion, da er eine Verringerung des e-Transport über den Cytochrom b6f bewirkt und die Abgabe überschüssiger Energie als Wärme (qE) auslöst, die Hauptkomponente des nicht-photochemischen Quenchings (NPQ). Pflanzen halten eine Balance zwischen Lichtschutz und Biomassegewinn mittels Feinabstimmung der PMF-Partitionierung. Ionentransportproteine in der Thylakoidmembran spielen dabei eine Schlüsselrolle, da der H+-Transfer in das Lumen häufig an ein Gegenion gekoppelt ist. Dieser Fluss dissipiert das Membranpotential und ermöglicht daher die H+-Akkumulation im Lumen. Es folgt daher, dass eine genaue Kenntnis des Thylakoid-Ionentransportoms eine Voraussetzung für unser Verständnis der PMF sowie für jegliche Bemühungen, photosynthetische Produktivität zu verbessern darstellt. In der Modellpflanze Arabidopsis thaliana sind Transportproteine, die den Cl-Fluss (VCCN1) und den K+/H+ Austausch (KEA3) vermitteln und daher Einfluss auf die PMF-Partitionierung haben bekannt. Mindestens ein zusätzlicher Transporter für K+ bzw. einwertige Kationen wird in der Thylakoidmembran erwartet. Projekt 6 ist auf NHD1 fokussiert, einen mutmaßlichen Na+/H+-Austauscher in der Thylakoidmembran. Die heterologe Expression von NHD1 stellt das Wachstum von E. coli-Mutanten wieder her, die im K+-Transport defekt sind. Dies deutet darauf hin, dass NHD1 auch den K+-Fluss über die Thylakoidmembran in vivo vermitteln könnte. Projekt 6 hat zum Ziel die Folgen von Verlust bzw. zusätzlicher NHD1-Funktion auf die PMF, die Photosynthese und die globale Genexpression in Arabidopsis thaliana zu untersuchen, Substratspezifität von NHD1 zu charakterisieren und höhere Mutanten ohne NHD1, VCCN1 und KEA3 zu designen und zu analysieren. Die gesammelten Daten werden eine Aktualisierung des Modells der PMF-Dynamik und ihrer molekularen Komponenten unter fluktuierenden Lichtbedingungen ermöglichen.
Mitochondrien und Chloroplasten haben das eukaryotische Leben maßgeblich geprägt. Im Zentrum der Funktion beider Organellen liegt die protonenmotorische Kraft (PMF). Die PMF verbindet die Redoxchemie des Elektronentransports mit der ATP Synthese und fungiert darüber hinaus als Regulationsknotenpunkt, der die Funktion und die Zusammensetzung beider Organellen massgeblich beeinflusst. In Pflanzen und Algen sind die mitochondrielle und die chloroplastidäre PMF über das zelluläre metabolische Netzwerk miteinander verknüpft. Obwohl die bioenergetischen Prozesse der Chloroplasten und der Mitochondrien aufeinander abgestimmt sind um den effizienten Betrieb der Photosynthese zu gewährleisten, ist es noch immer weitgehend unklar wie genau dieses Zusammenspiel in vivo erreicht wird und welche Proteine hierzu auf Ebene der PMF in welchem Umfang beitragen. Projekt 7 geht der Frage nach wie die PMF von Mitochondrien und Chloroplasten in vivo durch die subzelluläre Dynamik von Protonen (H+), Kaliumionen (K+) und Calciumionen (Ca2+) kontrolliert wird. Ein ausgewachsenes Rosettenblatt von Arabidopsis thaliana dient Projekt 7 als Modellsystem, um unter Verwendung von fluoreszierenden Protein-Biosensoren und hochauflösender in vivo Mikroskopie einen Atlas der Dynamik von H+, K+ and Ca2+ in den Subkompartimenten von Chloroplasten und Mitochondrien in folio zu erstellen. Neben dreidimensionaler molekularer Information in lebenden Blattgeweben wird dieser Atlas auch die physiologischen Antworten auf unterschiedliche Beleuchtungsregimes umfassen. Der Atlas wird so eine bislang einzigartige Referenz darstellen, die es ermöglicht zu verstehen welche Rolle Kandidatenproteine für den Transport von H+, K+ und Ca2+ über die organellären Membranen in der Festlegung der PMF Charakteristika in folio spielen. Projekt 7 testet eine Auswahl an genetischen Mutanten mit funktionellen Defekten in den Dynamiken von H+, K+ und Ca2+ in den Mitochondrien und den Chloroplasten auf deren Einfluss auf die organellären PMF Charakeristika in folio. Diese Kandidatenproteine umfassen KEA3 (gemeinsam mit Projekt 5) und NHD1 (gemeinsam mit Projekt 6) genauso wie MCUs, MICU, UCP1 und AOX1a. In Kombination werden das Atlaskonzept und die Mutantenanalyse Aufschluss über die zentrale Frage geben, in welchem Maße sich die PMF Charakteristika in einem Organell auf die des anderen Organells auswirken. Die Bedeutung einer veränderten PMF Dynamik für den Photosynthesestoffwechsel, die Ultrastruktur der Organellen und die Leistungsfähigkeit der gesamten Pflanze wird sich erschließen. Daraus wird sich ein erweitertes Verständnis der PMF Regulation in Chloroplasten und Mitochondrien und der Beiträge, die ausgewählte Proteine bei der Justierung der PMF Charakteristika leisten, ergeben.
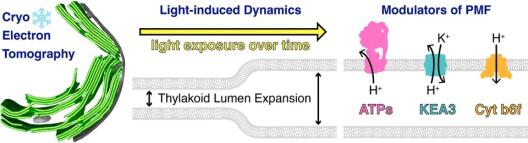
Thylakoidmembranen enthalten photosynthetische Komplexe, die den lichtgetriebenen Elektronentransfer mit der Ausbildung einer protonenmotorischen Kraft (PMF, bestehend aus ΔpH und ΔΨ) koppeln, welche die Produktion von ATP antreibt. Somit ist die Thylakoidarchitektur eng mit einer effizienten Photosynthese verbunden. Thylakoide sind flach statt kugelförmig, wodurch die Konzentration von photosynthetischen Komplexen pro Lumenbereich erhöht ist, während gleichzeitig eine Ausrichtung für die Diffusion des löslichen Elektronenträgers Plastocyanin durch das Thylakoidlumen entsteht. Die Thylakoidarchitektur ist jedoch nicht statisch, sondern reagiert auf Änderungen der Lichtqualität und -quantität. Vorläufige elektronenmikroskopische Beobachtungen in Cyanobakterien, Grünalgen und Pflanzen zeigen, dass das Thylakoidlumen unter erhöhter Lichtintensität anschwillt. Eine logische Hypothese wäre, dass die Ansäuerung des Lumens (ΔpH) und ionenflussgetriebene Änderungen des Membranpotentials (ΔΨ) mit osmotischen Kräften gekoppelt sein könnten, die das Lumen erweitern. Die genaue Dynamik und der detaillierte Mechanismus dieser architektonischen Veränderung sind jedoch unklar. Projekt 8 hat zum Ziel, die mechanistischen Grundlagen und physiologischen Folgen der lichtinduzierten Thylakoidschwellung zu verstehen und gezielt zu testen, wie diese Schwellung durch verschiedene Komponenten der PMF moduliert wird. Dies wird durch einen neu entwickelten Kryo-Elektronentomographie (Kryo-ET)-Workflow unterstützt, der es ermöglicht die native Thylakoid-Architektur (Membran- und Lumenbreite) mit Sub-Nanometer-Präzision zu messen und die Organisation jedes photosynthetischen Komplexes entlang dieser Membranen zu analysieren. Projekt 8 zielt zunächst darauf ab die Kinetik der Thylakoidschwellung in Grünalgen (Chlamydomonas reinhardtii) und Cyanobakterien (Synechocystis sp. 6803) zu charakterisieren, durch die Erfassung einer detaillierten Kryo-ET-Zeitreihe von Wildtypzellen nach dem Übergang von Dunkelheit zu verschiedenen Lichtintensitäten. Als nächstes beabsichtig Projekt 8 die Beiträge von ΔpH und ΔΨ zur Thylakoidlumenexpansion zu untersuchen, durch die Kombination von Kryo-ET mit kleinen Molekül-Ionophoren mit genetischen Störungen von ATP-Synthase (gemeinsam mit Projekt 4) und KEA3-Ionen-Antiportern (gemeinsam mit Projekt 5). Parallel dazu ist Projekt 8 bestrebt die spezifischen PMF-Beiträge des Cytochrom b6f Komplexes zur Dynamik des Thylakoidlumens zu testen (gemeinsam mit Projekt 1). Insgesamt zielt Projekt 8 darauf ab, ein integriertes Modell der lichtabhängigen Lumenexpansion zu erstellen, durch eine Kombination von Kryo-ET-Daten mit funktionellen Messungen der PMF und photosynthetischer Funktion.
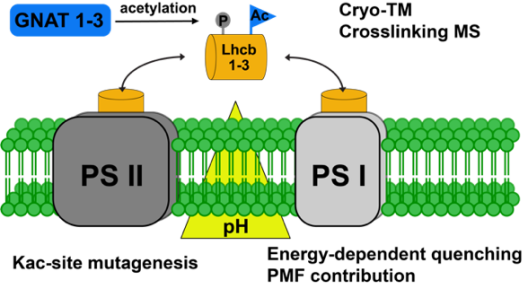
Der photosynthetische Elektronentransport ist zur Vermeidung von Photoinhibition und Schäden an den Photosystemen in Abhängigkeit von den Umweltbedingungen dynamisch reguliert. Insbesondere während der Akklimatisierung an veränderte Umweltbedingungen spielen chemische Proteinmodifikationen eine wichtige Rolle bei der Regulation des photosynthetischen Ertrags. Neben der Phosphorylierung ist die Acetylierung von Aminogruppen innerhalb von Proteinen eine wichtige posttranslationale Modifikation, die besonders häufig bei an der Photosynthese beteiligten Proteinen auftritt. N-terminale Acetyltransferasen oder Lysin-Acetyltransferasen katalysieren den Transfer der Acetylgruppe von Acetyl-Coenzym A auf die Ziel-Aminogruppe. Im Gegensatz zur N-terminalen Acetylierung ist die Acetylierung von Lysinresten (KAc) reversibel, wobei Lysin-Deacetylasen für die Entfernung von Acetylgruppen von Proteinen verantwortlich sind. Kürzlich wurden acht neue dualspezifische Acetyltransferasen (GNATs) in Chloroplasten von Arabidopsis thaliana identifiziert. Die erste Charakterisierung von GNAT2 zeigte, dass dieses Enzym für photosynthetische State Transitions unverzichtbar ist. Darüber hinaus zeigen die Mutanten im Vergleich zum Wildtyp ein erhöhtes energieabhängiges Quenchen bei hohen Lichtintensitäten. Obwohl die ersten Proteinsubstrate von GNAT2 identifiziert wurden, ist die spezifische Rolle der Proteinacetylierung bei der Akklimatisierung der photosynthetischen Lichtreaktionen auf mechanistischer Ebene noch nicht verstanden. Projekt 9 hat zum Ziel neue biochemische Einblicke in die Funktionen der Acetylierung ausgewählter Zielproteine zu geben und die Rolle der plastidären Acetyltransferasen GNAT1-3 bei der Regulation der Acetylierung am nicht-photochemischen Quenchen beteiligter Proteine zu untersuchen. Projekt 9 verwendet neueste gel-basierte chemische Crosslinking Techniken in Kombination mit Kryo-Elektronentomographie (gemeinsam mit Projekt 8) zur 3D-Modellierung um Unterschiede in der Zusammensetzung der Thylakoidproteinkomplexe in Abhängigkeit von verfügbarem Licht und Genotyp zu untersuchen. Projekt 9 ist insbesondere auf die Rolle der Acetylierung bei photosynthetischen Akklimatisierungsreaktionen durch Änderungen der Lichtverfügbarkeit fokussiert.
Die protonenmotorische Kraft (PMF) über die Thylakoidmembran spielt eine zentrale Rolle bei der Regulierung des Energiestoffwechsels von photosynthetischen Organismen. Die Zahl der voneinander abhängigen Prozesse und möglichen Querverbindungen zwischen Reaktionen, die PMF erzeugen, modulieren und von ihr abhängen, ist bemerkenswert, und ihre kombinierte Wirkung auf die Bioenergetik der Zelle führt zu einer nichtlinearen Komplexität. Daher ist ein strenger Rahmen erforderlich, der eine systematische Entflechtung dieser Komplexität ermöglicht, um Konzepte zu entwickeln, wie die PMF in verschiedenen photosynthetischen Arten erreicht und reguliert wird. Computergestützte, mechanistische Modelle der photosynthetischen Elektronentransportkette bieten einen quantitativen Rahmen, der vergleichende Studien ermöglicht. Zahlreiche Modelle der Photosynthese beschreiben die PMF, aber viele von ihnen konzentrieren sich ausschließlich auf ihre pH-abhängige Komponente (ΔpH), vor allem wegen ihrer Verbindung zu dem wichtigen photoprotektiven Mechanismus, dem nicht-photochemischen Quenching. Beim energieabhängigen Quenching wird überschüssige Lichtenergie als Wärme freigesetzt, um das Photosystem II vor Schäden zu schützen. Die Bedeutung des Membranpotenzials (ΔΨ) für die Erzeugung der PMF und die Photoprotektion sollte jedoch nicht außer Acht gelassen werden, da der photosynthetische Elektronentransport sehr empfindlich auf die Konzentration von Ionen (z. B. H+, K+, Mg2+ und Cl-) reagiert. Ionentransportproteine in der Thylakoidmembran, wie z. B. KEA3, ermöglichen einen Ausfluss von H+ mit einem Gegenfluss von Kationen, wodurch der Beitrag der protonenbasierten oder potenzialgetriebenen Komponenten der PMF beeinflusst wird. Durch eine dynamische Feinabstimmung der beiden PMF-Komponenten können sich photosynthetische Organismen dynamisch an Umweltveränderungen anpassen und ihre Energieproduktion auf ihren Energiebedarf abstimmen. Projekt 10 hat zum Ziel einen vereinheitlichenden theoretischen Rahmen zu schaffen, der die Dynamik der PMF in zahlreichen Klassen photosynthetischer Organismen beschreibt, die im Rahmen von GoPMF untersucht werden, von Cyanobakterien über grüne Mikroalgen bis hin zu vaskulären Pflanzen. Aufbauend auf zuvor erstellten mathematischen Modellen der photosynthetischen Elektronentransportkette erforscht Projekt 10 den modularen Aufbau dieser Modelle, um einen Bauplan für die PMF-Regulierung zu erstellen. Innerhalb dieses Rahmens untersucht Projekt 10 systematisch die Auswirkungen der PMF-Aufteilung auf die photosynthetische Dynamik und stellt speziesübergreifende Vergleiche her um neue Hypothesen zur PMF-Regulierung zu entwickeln. In Verbindung mit den gesammelten experimentellen Arbeiten von GoPMF wird das Modell darüber hinaus die Experimente zur lichtgetriebenen Wasserstoffproduktion in Cyanobakterien leiten.
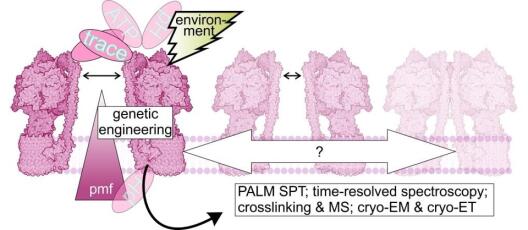
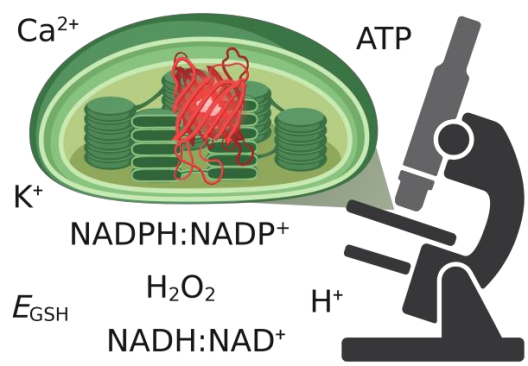
Die biologische Forschung hat durch rasche Entwicklungen in der hochauflösenden Bildgebung und der in vivo Biosensorik rasante Fortschritte erzielen können. Eine Kombination beider Ansätze erlaubt die in situ Quantifizierung zellphysiologischer Parameter und die Visualisierung von (sub)-organellen und molekularen Dynamiken. Diese Entwicklungen haben unser Verständnis der protonenmotorischen Kraft (PMF) revolutioniert. Die Visualisierung von pH-Dynamiken und Δψ in Mitochondrien hat ein unerwartetes Bild der PMF-Organisation und ihrer dynamischen Veränderungen in einzelnen Organellen gezeichnet. Um ein vergleichbares Verständnis der PMF über photosynthetische Membranen zu etablieren, ist die Entwicklung neuer spezieller Methoden erforderlich. Ein umfassendes Verständnis der PMF in vivo benötigt auch die Etablierung von Sensoren für Ionen wie K+, die zur PMF-Partitionierung beitragen, und zugehörige Metaboliten wie NADPH:NADP+, NADH:NAD+ und ATP, die eng mit der chloroplastidären PMF verknüpft sind. Da viele der jetzt verfügbaren Biosensoren proteinbasiert sind, ermöglichen spezifisches subzelluläres Targeting und genetische Fusion mit Proteinen in bioenergetischen Membranen Messungen mit höchster lokaler Spezifität. Dafür ist jedoch eine umfassende Charakterisierung der biochemischen und biophysikalischen Eigenschaften der Biosensoren erforderlich, sowohl isoliert als auch in situ, um ihre Spezifität sicherzustellen und Artefakte zu vermeiden. Spezialisiertes technisches Wissen stellt hierbei eine Voraussetzung dar, die realistischerweise nicht alle individuellen Forschenden erfüllen können. Das Zentralprojekt führt das verfügbare Fachwissen zur Biosensorik mit neuen experimentellen Erkenntnissen zusammen. Darüber hinaus entwickelt das Zentralprojekt Biosensor-Strategien für potosynthetische PMF-Messungen und optimiert hochauflösende Lichtmikroskopie von Chloroplasten sowie photosynthetischen Membranen. Insbesondere etabliert das Zentralprojekt die stromale und lumenale Biosensorik von H+ Ionen um den ΔpH direkt auszulesen. Zudem leistet das Zentralprojekt über NADPH:NADP+-Biosensorik Pionierarbeit für zukünftige Projekte und optimiert maßgeschneiderte Setups zur Biosensorik unter veränderlichen photosynthetischen Bedingungen. Das Zentralprojekt hat zum Ziel, superauflösende Bildgebung mit sub-organellarer Auflösung für potosynthetische Membranen zu entwickeln. Dies ist nicht auf Live-Bildgebung durch konfokale Mikroskopie mit modernster Dekonvolution und Airyscan-Detektion beschränkt, sondern umfasst auch die Entwicklung fortschrittlicher Protokolle zur Proteinmarkierung für hochauflösende Bildgebung, Einzelpartikellokalisierungsmikroskopie und Mobilitätsstudien von Thylakoidmembranproteinen. In enger Zusammenarbeit mit den Mitgliedern und Projekten von GoPMF erweitert, verfeinert und implementiert das Zentralprojekt kontinuierlich die Toolbox zur optischen Analyse von PMF-Determinanten.