|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Free Neuropathology 5:35 (2024) |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Review |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Cryopreservation of brain cell structure: a review |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Andrew T. McKenzie1, Emma L. Thorn2,3, Oge Nnadi4, Borys Wróbel5,6, Emil Kendziorra7, Kurt Farrell2,3, John F. Crary2,3 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Corresponding authors: |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Additional resources and electronic supplementary material: supplementary material |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Submitted: 11 September 2024 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Keywords: Cryopreservation, Vitrification, Brain banking, Cryoprotectant, Cryofixation, Ice artifact |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Abstract Cryopreservation, the preservation of tissues at subzero temperatures, is a mainstay of brain banking that allows for the storage of brain tissue without the use of chemical fixatives. This is particularly important for molecular studies that are incompatible with tissue fixation. However, brain tissue is vulnerable to various forms of damage during the cryopreservation process, in particular due to the phase transition of water from a liquid to a solid state with the formation of ice crystals, which can disrupt cellular morphology. There is a critical need to characterize the effects of cryopreservation on brain cell structure at the microscopic level. In this review, we conducted a comprehensive literature search, identifying 97 studies that yielded 146 distinct observations of the effects of cryopreservation on neurohistology. We classified the reviewed studies into three main categories: cryofixation, freezing, and cryopreservation with cryoprotectants. Cryofixation techniques enable vitrification and excellent ultrastructural preservation of thin tissue samples but are limited in terms of the depth of tissue that can be preserved without ice artifacts. Freezing methods, particularly when applied to brain slices, can achieve rapid cooling rates that result in minimal ice artifacts detectable by light microscopy. Cryoprotectant-based approaches have the potential to reduce ice damage and achieve vitrification. For thin tissue samples, immersion in cryoprotectants has been found to be effective for structural preservation. However, for larger samples or the entire brain, perfusion of cryoprotectants is necessary to perform rapid distribution, and this has a more limited evidence base. In conclusion, while current cryopreservation methods can provide sufficient quality for some downstream applications, there is a need for improved techniques that enable the cryopreservation of larger brain tissue samples while maintaining excellent structural preservation. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
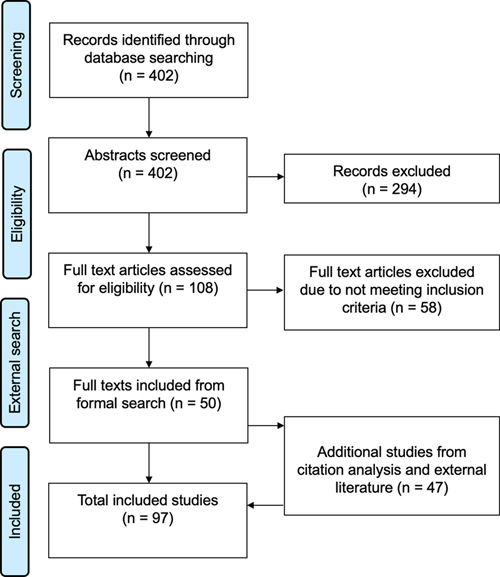
Introduction Cryopreservation is a key technique in contemporary brain banking (McKee, 1999; Vonsattel et al., 2008; Ramirez et al., 2018). Here, we define cryopreservation as the preservation of tissue at temperatures below 0 °C. This can allow for the long-term storage of preserved brain tissue for subsequent research, where it is frequently used for biomolecular profiling studies such as nucleic acid sequencing or metabolomics. Cryopreservation preserves tissue by decreasing the rate of molecular motion across an entire biospecimen (Wowk, 2010; Weng et al., 2019). However, cryopreservation can cause several types of structural artifacts, including ice formation, dehydration, displacement of tissue structures, and local aggregation of biomolecules (Rosene and Rhodes, 1990; Jain et al., 2021). These artifacts can be ameliorated by vitrification, wherein tissue is cooled to form a glass-like amorphous solid without ice crystal formation, although this can be difficult to achieve in the large tissue volumes typical of brain banking (Wowk, 2010). Advances in refrigeration, cryogen production, cryobiology, and their application in brain banking over the last several decades have allowed a set of best practices to emerge for how brain tissue should be cryopreserved. For example, many brain banks rapidly cool slabs of fresh brain tissue between cooled metal plates (Jones et al., 1992; Vonsattel et al., 2008). Despite these advances, better cryopreservation solutions are clearly needed for many potential research applications. As a result, there is a critical need to examine the effects of cryopreservation on brain cell structure, to better understand what the state-of-the-art methods are and how they could potentially be improved upon. Our review focuses on unfixed tissue. However, we note that there is an extensive literature showing that brain tissue, which is pre-fixed with chemicals can be effectively cryopreserved with the use of cryoprotectants (Watson et al., 1986; Meissner and Schwarz, 1990; Otubo et al., 2021). Chemical fixation aids in the process of cryoprotection because it prevents decomposition during cryoprotectant diffusion, it mitigates damage due to dehydration, and it stabilizes biomolecules in place (Paljärvi et al., 1979; Robards and Sleytr, 1985, p. 88; Meissner and Schwarz, 1990). Perfusion with fixatives has also been shown to help with subsequent perfusion of cryoprotectant, possibly because fixation helps to stabilize blood vessels (McIntyre and Fahy, 2015). With the use of sufficient cryoprotection, the tissue can be vitrified, entirely preventing ice formation and associated ice artifacts (McIntyre and Fahy, 2015). Chemical fixation prevents cellular damage during cryoprotection, aids with the membrane permeability of cryoprotectants, and may help stabilize membrane structure by “hardening” the lipids (Moor et al., 1980). There are methods available in practical brain banking settings for fixation followed by cryopreservation and long-term storage (Estrada et al., 2017; Kumarasami et al., 2023; Gasperoni and Dworkin, 2024). However, for some research applications, fresh tissue is preferable to fixed tissue. Therefore, in this review, we focus on the cryopreservation of unfixed brain tissue. In this review, we examine existing data on the cryopreservation of brain tissue, identifying previous methods used and their impacts on histologic integrity. In the studies we identified, the most common long-term storage temperatures range from ‑20 °C to ‑196 °C (liquid nitrogen temperature). We include approaches that attempt to freeze tissue while tolerating some degree of ice formation as well as approaches that attempt to vitrify tissue and thereby avoid ice formation. We focus on the effects of cryopreservation on brain structure as identified via microscopy, rather than the features of brain tissue identified by neuroimaging, electrophysiology, cell culture, or dissociated biomolecular profiling. We discuss the mechanisms of tissue damage in cryopreservation, the efficacy of different cooling methods, the use of chemical cryoprotectants to decrease ice crystal formation, and other factors that affect variability in cryopreservation outcomes between studies. Through this review, we aim to illuminate the current landscape of cryopreservation techniques, identify gaps in knowledge, and discuss potential avenues for improvements. This review may be useful for individuals banking brain tissue via cryopreservation, by offering a summary of the upsides and downsides of the different methods available. In addition, researchers who analyze cryopreserved brain tissue may find this review valuable, as it summarizes our current knowledge about the expected burden of freezing artifacts, as well as which downstream research applications may be the most compatible with cryopreserved brain tissue. Methods Realist synthesis We carried out a realist synthesis review aimed at establishing a theoretical understanding of cryopreservation. This method integrates elements of a systematic review with an emphasis on theory and practical application (Wong et al., 2013). We chose this approach due to the wide-ranging methods used in cryopreservation of brain tissue. We report our adherence to the RAMESES standards (Supplementary File 1) (Wong et al., 2013). Before formalizing our search methodology, we first scoped the literature by ad hoc searches of PubMed, Google, Google Scholar, and by holding discussions among the authors. The review protocol was preregistered here: https://osf.io/y6hv2. Eligibility criteria To be included in this review, studies needed to report original empirical data relevant to the effects of unfixed brain tissue cryopreservation on histologic integrity. We included studies that met the following two criteria: (1) Examined the cryopreservation of unfixed brain or other nervous system tissue. Isolated cells, sub-cellular structures, clusters of cells, or organoids alone were not sufficient. (2) Assessed histological and/or morphological preservation through microscopy or other structural analysis methods. Through this criterion, we focused on structural and morphological integrity rather than functional or biomolecular preservation alone. Both human and non-human animal studies were eligible for inclusion. If multiple publications report on the same underlying data, we only included the study that drew the most relevant conclusions about the effects on cryopreservation. When a single study reported multiple independent observations about the effects of cryopreservation, such as a different type of brain sample, cooling rate, or rewarming method, they were each extracted from the study as separate data points. Search methods and study selection We searched PubMed to identify relevant studies with keywords related to cryopreservation of brain tissue. We searched the PubMed database from its inception to April 18th, 2024, using the following search query: «(freezing[tiab] OR ice[tiab] OR cryopreservation[tiab] OR "Cryopreservation"[Mesh]) AND (histology[tiab] OR microscopy[tiab] OR ultrastructure[tiab] OR "electron microscopy"[tiab] OR "ultrastructure"[Subheading]) AND (brain[tiab] OR "central nervous system"[tiab] OR "Brain"[Mesh])». One reviewer screened the titles and abstracts and identified studies for full-text review based on the eligibility criteria (Supplementary File 2). Next, one reviewer evaluated the full texts and selected studies to include (Supplementary File 3). To find additional eligible studies, we also (a) hand-searched the reference lists and referencing studies of the most relevant papers and (b) performed additional ad hoc searches of the literature relevant to this topic. Any studies meeting the inclusion criteria that were identified outside the formal search process, such as in the initial literature scoping process, were also included. Data extraction For each included study, we extracted relevant data: species; specimen type; cryoprotectant, if any reported; cooling method; storage temperature; storage length; rewarming method; visualization method; any additional outcome specifier reported; the histological or morphological outcome(s) described; and any relevant quote or quotes. We treated each distinct cryopreservation condition, tissue type, visualization method, or other methodologic difference reported in a study as a separate “observation”. For example, if a study reported outcomes for tissue cooled at two different rates, these were recorded as two observations. However, minor variations in otherwise highly similar cryopreservation conditions or visualization methods from the same study were grouped into a single observation. The determination of whether two similar cryopreservation conditions or visualization methods should be grouped into one observation or separated was based on an assessment of the level of detail and independence reported for each in the text. For each study, one reviewer performed the data extraction. On randomly selected articles, chosen via https://www.random.org/, one other reviewer then evaluated the extracted data. This independent evaluation process was performed on n = 20 (21 %) of the studies and did not identify any major discrepancies in the extracted data. The extracted data table is available here: https://github.com/andymckenzie/Cryopreservation_Review. Qualitative data analysis Due to the wide range of cryopreservation techniques and outcomes reported, we performed a qualitative synthesis rather than attempting formal quantitative meta-analysis or grading of outcomes. We drew insights from the extracted data to categorize studies and identify key themes. We categorized the included studies into three partially overlapping types based on the cryopreservation approach used: (1) cryofixation, (2) freezing, and (3) cryopreservation with cryoprotectants. We focused on comparing histologic outcomes between different methods of cryopreserving brain tissue, as well as identifying mechanistic insights into the causes of these outcomes. Example frozen brain tissue data The brain tissue used was obtained and de-identified at the Icahn School of Medicine at Mount Sinai in accordance with its policies, regulations, and institutional review board recommendations. All brain tissue processing was performed in the Neuropathology Brain Bank & Research CoRE. The brain donor was female and was 76 years old at the time of death. The postmortem interval from time of death to freezing was 70.5 hours. The brain was removed from the skull and placed in a cooler for transport from the autopsy suite to the brain bank. The brain was bisected into two hemispheres. The right hemisphere was immersion fixed in toto in 10 % neutral buffered formalin, while the left hemisphere was sectioned fresh into approximately 0.5 cm coronal slices. These fresh slices were parcellated into approximately 2 x 2 cm blocks, and then cryopreserved between Teflon-coated metal plates cooled at ‑80 °C. The tissue was stored at ‑80 °C for 5.3 years. One frozen tissue block of the lateral occipital cortex was then moved to a ‑20 °C freezer for 6 hours and 10 minutes. Formalin fixative was also frozen at ‑20 °C and then moved to 4 °C at the same time as the tissue. The tissue was placed in the slushy consistency formalin at 4 °C for 18 hours before being moved to phosphate-buffered saline (PBS) containing 0.1 % sodium azide at room temperature for 4 hours and 45 minutes. It was then placed in room temperature formalin for 55 minutes before being processed for histology. Frozen-thawed tissue from the lateral occipital cortex region was compared to immersion fixed tissue from the calcarine cortex of the contralateral hemisphere, which was fixed in toto for 37 days. The fixed tissue was paraffin embedded, and a microtome was used to prepare consecutive sections with a thickness of 5 µm, as previously described (McKenzie et al., 2022). These sections were then placed on glass slides, deparaffinized, and stained with hematoxylin and eosin, followed by imaging with a digital slide scanner. Characteristics of included studies Through our formal search procedure, we screened 402 abstracts, assessed 108 full-text articles, and identified 50 studies meeting our inclusion criteria (Figure 1). An additional 47 studies were included based on citation analysis or supplementary ad hoc searches, yielding a total of 97 studies for this review. We categorized the included studies into three partially overlapping types based on the cryopreservation approach: (1) Cryofixation studies, which used ultrarapid cooling methods like high-pressure freezing or plunge freezing to achieve vitrification of at least part of the tissue; (2) Freezing studies, which cooled tissue without cryoprotectants at various rates, allowing some degree of ice crystal formation; and (3) Cryopreservation with cryoprotectants studies, which used substances like DMSO or glycerol to reduce ice crystal formation during the cooling process. The studies were performed on tissue from various species, including humans, rats, and mice. Before discussing the outcomes reported in these studies, we first review the mechanistic aspects of cryopreservation that provide important context for interpreting the results.
Figure 1. Study selection flow diagram. Mechanisms of brain cryopreservation Physical aspects of cooling To understand the effects of cryopreservation on brain cell structure, it is helpful to build up a basic understanding of what happens during cryopreservation of brain tissue (McGrath, 1985; Karlsson and Toner, 1996; Wowk, 2010). Note that this section is not meant to be a comprehensive summary of this complex topic, but rather to provide a context for the observed phenomena and outcomes in the empirical studies we reviewed. The reason that cooling can act as a preservation method is that as temperature decreases, the kinetic energy of molecules is reduced, leading to slower molecular motion and chemical reactions. At low enough temperatures, a specimen, or parts of a specimen, will convert from a viscous liquid to a glass-like state, in the process of vitrification (Fahy and Wowk, 2021). This effectively “pauses” biological processes, including those that lead to cell death and tissue decay (Wowk, 2010). However, as the temperature decreases below the freezing point of water, water molecules also have a thermodynamic tendency to spontaneously rearrange into ice crystals, which cause damage to the tissue. In biological tissues, there is a further complication, which is the potential for non-water substances to crystallize, such as in the process of eutectic crystallization. We will discuss the effects of ice formation first. How ice damages biological tissue Ice formation leads to damage to brain cell structure through several mechanisms. These forms of damage can collectivity be called “freezing injury”. First, formed ice crystals can cause mechanical damage to local structures as they expand, such as causing the displacement or rupture of cell and organelle membranes (Fahy and Wowk, 2021). Second, as water is drawn towards growing ice crystals, this causes large stresses to build up near the ice-water interface, which is a phenomenon known as cryosuction (Gerber et al., 2022). This stress is associated with high local pressures of around 1 MPa per degree of temperature below the freezing point, leading to potential alterations of biomolecular distributions in the local area. Third, as extracellular and intracellular water is removed and cells become dehydrated, high solute concentrations in the cells lead to structural damage (Fahy and Wowk, 2015). Damage due to high solute concentrations has been called “solution effects” (Mazur et al., 1972). For example, solution effects have been reported to cause the denaturation and aggregation of biomolecules (Li et al., 2023). The formation of ice during freezing can be used to permeabilize cells, which has been attributed to both mechanical disruption of membranes by ice and osmotic forces (Zhang et al., 2016). Smaller ice crystals may be particularly likely to cause the membrane damage that contributes to cell permeabilization (Zhang et al., 2016). As cell membranes are permeabilized, this allows soluble biomolecules to leak out of cells and potentially diffuse away from their original locations, which is another form of damage to brain cell structure during cryopreservation (Rossner et al., 2006; Notter et al., 2014; Gellhaar et al., 2017). The location of ice formation across tissues is not random, but rather can be predicted to some degree. In many organs, ice formation tends to propagate along the vascular system, likely because there is no barrier to crystal growth in these areas (Rubinsky and Pegg, 1988). When these tissues are examined under a microscope, the structural changes caused by large ice crystal formation also follow predictable patterns. Upon rewarming, areas of tissue that previously underwent ice formation tend to have anisotropic spaces empty of tissue, indicative of the crystalline structures of ice when visualized under a microscope. These ice damage artifacts have been described as vacuoles, “Swiss cheese holes,” or as having a “sponge-like” appearance (Chambers et al., 1932; Taqi et al., 2018). In addition to generating open spaces, ice formation can cause other structural changes, such as disrupting membrane structures or creating a web-like pattern (reticulation) in the nucleus and cytoplasm (Möbius et al., 2010). This reticulation can look like irregularly shaped dark strands when viewed under an electron microscope. After severe ice damage, the tissue can be dramatically modified. It has been reported that large ice crystals can distort cell membranes and organelles to such an extent that the pre-existing structural configuration is unrecognizable and the underlying structure of the specimen appears to be destroyed (Ryan, 1991). How ice damage can be prevented One method available to entirely prevent ice formation is the use of an extremely rapid cooling rate. If the temperature of the biospecimen is reduced very quickly, ice does not have the time to form or grow to a meaningful degree. The minimum rate of cooling necessary to cool a sample fast enough to “outrun” the speed of ice formation and achieve vitrification is called the critical cooling rate (Fahy and Wowk, 2021). Because the required cooling rate is very rapid, this generally requires exposing very small samples of cells or tissues to extremely cold temperatures. In larger samples, it is not possible to perform this, because the slow rate of dissipation of heat by convection in biological tissue prevents it. For example, one source reports that an infinitely fast cooling rate at the surface of an aqueous sample would lead to a cooling rate of around 5000 K/s at the depth of 100 µm, which is not sufficiently fast for vitrification (Studer et al., 2014). Another method to prevent ice formation is to add cryoprotectants to the tissue. Cryoprotectants are chemicals that decrease or prevent freezing injury during cryopreservation (Fahy and Wowk, 2015). Cryoprotectants can aid in vitrification by directly forming hydrogen bonds with water molecules (Lee and Baiz, 2022), causing freezing point depression as a colligative effect and/or increasing the viscosity of the tissue. Although there are endogenous chemicals in tissues that act as cryoprotectants, the term “cryoprotectant” usually refers to an external chemical that is added to a tissue for this purpose. These substances often replace water as the tissue solvent. Examples of cryoprotectants include small molecules such as glycerol, ethylene glycol, dimethyl sulfoxide (DMSO), or ice blockers such as antifreeze proteins (Fahy and Wowk, 2015). Notably, cryoprotectants and faster cooling rates can work together to protect a tissue from ice damage during cryopreservation. With a higher concentration of cryoprotectants, a lower cooling rate is necessary to prevent ice damage, and vice versa (Wowk, 2010). By preventing ice formation, both cryoprotectants and faster cooling rates are expected to improve structural preservation quality during cryopreservation. However, the introduction of cryoprotectants can also alter cellular and molecular structures (Robards and Sleytr, 1985, p. 77). Most commonly, the high concentration of cryoprotectants needed to prevent ice formation leads to cell shrinkage and/or osmotic stress, particularly if the addition of cryoprotectants is not performed in a graded manner. To minimize damage from osmotic stress, cryoprotectants are typically added slowly, allowing cells time to equilibrate and adjust to the changing osmotic environment. In larger samples, the slow addition of cryoprotectants via immersion also renders the tissue vulnerable to decomposition during this process. One way to minimize the time of cellular decomposition is to perfuse cryoprotectants through the blood vessels of the brain, which allows the cryoprotectants to extravasate into the tissue. However, perfusion of the postmortem brain is a challenging problem, especially in the setting of extended periods without blood flow (McFadden et al., 2019). Ice formation during freezing procedures In the absence of cryoprotectants, complete vitrification is not achievable for larger samples. Therefore, ice crystals will form. The total amount of ice formed is primarily dependent on the water content of the tissue and the final temperature reached, rather than on the cooling rate. For example, in one study of frozen sponge cake, the total ice volume fraction – about 60 % – was the same in samples frozen with fast (17.2 °C/min) and slow (0.3 °C/min) cooling rates (Zennoune et al., 2022). However, the distribution and size of ice crystals can differ substantially based on the cooling rate, which can have important effects on microstructural preservation. Relatively faster cooling results in the formation of numerous but smaller ice crystals, which are less disruptive to cellular structures (Pérez-Bermúdez et al., 2023). This is because rapid cooling reduces the time available for ice crystal growth and rearrangement. Smaller, more numerous ice crystals that form during rapid cooling are generally found to be less damaging to tissue morphology than larger crystals that form during slow cooling. Notably, sufficiently small ice crystals are not expected to be visible via light microscopy (Szabo and Burg, 2024). It is also helpful to understand mechanistically what occurs during slower cooling processes without a sufficient concentration of cryoprotectants to vitrify the entire tissue. In this situation, if the temperature is lowered to below the glass transition temperature, then vitrification is still expected to occur in localized areas between pockets of ice crystals. This is because as ice formation progresses during cooling, the concentration of solutes in the remaining unfrozen areas eventually becomes high enough to cause these areas to vitrify when the temperature reaches the glass transition temperature (Wowk, 2010). Localized vitrification within confined spaces during slow cooling is particularly relevant for intracellular spaces, which are densely packed with proteins and other biomolecules (Clarke et al., 2013). Cryoprotectants can protect cells from morphological damage during this process by limiting ice formation and preventing harmful, elevated salt concentrations. For example, in a study of slowly cooling Jurkat cells (an immortalized T cell line) in the presence of DMSO, an endothermic event was observed around ‑47 °C, which was attributed to a colloidal-like glass transition in the cytoplasm resulting from cellular dehydration and increased intracellular macromolecular crowding (Meneghel et al., 2019). The specific temperature of any intracellular glass transition will vary depending on many factors, such as cell type, protein content, solute concentration, and the concentration of any cryoprotectants. In the context of brain tissue, even if some degree of intracellular vitrification occurs, the fine cell processes or cell-cell connections may be mechanically damaged if extracellular ice is formed. Eutectic crystallization Although not commonly discussed, we note for completeness that eutectic crystallization can occur during cryopreservation in addition to the crystallization of water forming ice. A eutectic system is a mixture of substances that has a melting point below either of the original substances. When the melting point of this mixture is reached, the eutectic system can solidify. In practical terms, sodium chloride tends to be the main relevant electrolyte in biological tissues like the brain (Han and Bischof, 2004). As ice forms during cryopreservation, the water-sodium chloride solution increases in concentration and it can form a eutectic system because the mixture has a melting point lower than either of the individual components. In one study, eutectic crystallization of water-sodium chloride solutions – forming hydrohalite – occurred at approximately ‑30 °C, but this may differ based on the cooling rate (Han and Bischof, 2004). In yeast cells, hydrohalite has been found primarily around cell membranes (Okotrub and Surovtsev, 2013). Such eutectic crystallization can cause damage to cellular structures beyond that caused by ice formation alone (Han and Bischof, 2004; Klbik et al., 2022). It is unclear how much eutectic crystallization occurs in brain tissue during cryopreservation. However, it theoretically could pose additional challenges for maintaining cellular morphology. Notably, the use of cryoprotectants or the application of very rapid freezing techniques can potentially prevent eutectic crystallization. Cellular decomposition Another issue during cryopreservation is cellular decomposition by autolysis (self-digestion by cellular enzymes) and putrefaction (bacterial breakdown of tissues). These processes can occur during cooling and thawing of large brain tissue samples because they do not cool instantaneously, due to the limits of convective heat transfer (Pontén et al., 1973). Autolysis begins immediately after death when cellular enzymes are released, while putrefaction typically starts days later when bacteria proliferate. Importantly, cellular decomposition is dramatically inhibited by lower temperatures (Krassner et al., 2023). This is another reason that swift cooling is critical in cryopreservation protocols. Fractures One downside of cooling to very low temperatures is the possibility of fractures. Fracturing can occur in tissues or organs that are vitrified or frozen. It has been attributed to thermal gradients – i.e. differences in temperature in different parts of the organ (Woods and Mullen, 2016). It seems to be more common in larger specimens. One study found that brain tissue blocks of larger than 2.5 to 3.0 cm tend to fragment when immersed in liquid nitrogen-cooled liquid isopentane (Vonsattel et al., 2008). Fracturing has also been reported when whole rodent brains are cooled in liquid nitrogen (Peters, 2010). Fractures are more likely to occur at lower temperatures, as thermal stress accumulates. One study found that thermal stress in a cryoprotective solution is negligible until a temperature of around 5–10 °C above the glass transition temperature, depending on the viscosity of the solution (Rabin et al., 2006). Slower cooling rates allow more time for temperature to equilibrate across different areas of the organ or tissue, lowering the magnitude of thermal gradients. While slower cooling rates are associated with lower fracture rates, this effect appears to be relatively modest. For example, in one study, increasing the cooling rate by three orders of magnitude, from 0.5 °C/min to 500 °C/min, decreased the temperature at which thermal stress began to build up and fractures began to occur by only 7.2 °C (Rabin et al., 2006). The formation of brittle ice crystals in tissue can not only enhance fracturing during cooling or storage, but also during subsequent sectioning, posing a challenge for sample preparation of cryopreserved tissue (Marshall, 1940). Summary Most of the structural damage that occurs during cryopreservation of brain tissue is related to ice formation (Table 1). In addition, there are other forms of structural damage, such as eutectic crystallization, cellular decomposition, and fractures that can be independent of ice formation.
Cryofixation Preservation process Cryofixation refers to the ultrarapid cooling of tissue that is fast enough to prevent ice formation during cryopreservation (Gilkey and Staehelin, 1986). Major types include (a) plunge freezing, which directly immerses a sample into a liquid cryogen, (b) propane jet freezing, which sprays a liquid cryogen onto a sample, (c) cold metal block freezing, which rapidly brings the sample into contact with a pre-cooled metal surface, and (d) high-pressure freezing, which performs the cooling in a high pressure system to help suppress ice formation (Gilkey and Staehelin, 1986). One source reports that, in tissues without any cryoprotectants added, vitrification will result when it is cooled at a rate faster than 200,000 K/s (Studer et al., 2014). By increasing the pressure to around 2000 bar, high-pressure freezing lowers the freezing point of water to ‑22 °C, which reduces the critical cooling rate for ice formation by a factor of 100 (Studer et al., 2014). The vitrification depth of a cryofixation procedure is the distance from the surface of the tissue that can be vitrified. With the use of plunge freezing or cold metal block freezing, the vitrification depth is usually approximately 10–20 µm, but this can be increased 10-fold to samples around 200 µm thick with the use of high-pressure freezing (Studer et al., 2014). While increased pressure to this degree can be lethal, it can be kept to a very short period of less than 0.1 seconds during the high-pressure freezing procedure (Moor et al., 1980). As a result, the increased pressure is expected to have minimal effect on the tissue, although it has been reported to affect the structure of some biomolecules (Vanhecke et al., 2010). Occasionally, cryoprotectants are also used to aid in the process of cryofixation. This helps to prevent ice damage, but can lead to structural alterations of their own. As a result, cryoprotectants are only sometimes used in cryofixation, depending on the goals of the experiment. Notably, potential damage during rewarming is often avoided entirely for cryofixation studies because investigators frequently use freeze substitution, or less commonly freeze fracture, to avoid this issue. Effects on tissue morphology The majority of studies we identified found that cryofixation, and especially high-pressure freezing, can enable pristine ultrastructural preservation of brain tissue in the areas that are vitrified (Table 2; Supplementary File 4) (Benson et al., 1985; Guo et al., 2020). Cryofixation preserves membranes, organelles, synapses, and other neural structures with life-like morphology. Indeed, vitrification by cryofixation is considered the best method for structural preservation, superior to chemical fixation, because it can preserve tissue within milliseconds, rather than chemical fixation, which takes at least seconds to minutes (Sosinsky et al., 2008).
There is a large literature comparing vitrification via cryofixation to chemical fixation, which are two of the major ways currently available to ascertain native tissue structure. In general, most studies comparing the two methods have found that the ultrastructural appearance is largely similar, with cryofixation sometimes showing subtle improvements in the preservation of certain features (Landis and Reese, 1981, 1983; Reger and Escaig, 1988; Zhao et al., 2012; Korogod et al., 2015). For example, one study found that cryofixation of samples by placing them against a metal surface cooled to ‑196 °C better maintained the naturally smooth contour of cell processes and organelles (Hirokawa and Kirino, 1980). The authors also suggested that cryofixation appears to better preserve the extracellular spaces and better retain water-soluble components. Better maintenance of water-soluble components, may in turn lead to a more natural density of the cytoplasm and the mitochondrial matrix (Hirokawa and Kirino, 1980). It has been reported that microtubules are better preserved after cryofixation (Moor et al., 1980). Cryofixation also leads to better preservation of antigenicity (Hisano et al., 1986). Finally, cryofixation likely preserves a more natural distribution of water and ions between intracellular and extracellular compartments (Harreveld and Fifkova, 1975; Moor et al., 1980). However, despite these benefits, the differences between cryofixation and chemical fixation are generally subtle when both techniques are optimally performed. Indeed, tissue that is chemically fixed and cryoprotected with glycerol has been used as a control to ensure that the cryofixation procedure adequately preserves the structure of the tissue (Heuser et al., 1976). Many of the chemical fixation artifacts that have been described, such as cell shrinkage and membrane discontinuities, appear to be largely related to the tissue processing steps after fixation rather than due to the initial fixation itself (Sosinsky et al., 2008). Ice damage Cryofixation can enable excellent ultrastructural preservation in successfully vitrified regions. However, it is expected that in sufficiently deep areas of the tissue, where the cooling rate is insufficient to achieve vitrification, there will be ice crystal formation. The effects of ice crystals on tissue morphology manifest depending on the depth of the tissue and the size of the crystals. In the areas that border vitrified tissue, where the cooling rate becomes too slow to vitrify the tissue, small ice crystals and more subtle signs of suboptimal preservation are expected. For example, there may be blebbing of the nuclear envelope or alterations of the mitochondria (Möbius et al., 2010). Membranes may not show their clear expected structure (Senda et al., 2005). In deeper areas, the ice crystals are expected to be larger and to increasingly distort cellular structures, pressing the latter into one another, and causing large areas to be entirely devoid of structure (Heuser et al., 1976; Senda et al., 2005). Several studies also describe ice artifacts in deeper regions as causing reticulation of intracellular areas (Linner et al., 1986; Siksou et al., 2007; Möbius et al., 2010). Damage by tissue cutting and handling To isolate a sample that is sufficiently thin for cryofixation, cutting the tissue is generally required. This in turn leads to compression artifacts near the cut surface of the tissue, which can be severe (Zuber et al., 2005). One study omitted the first 20 µm of the sample from their analysis as a result of these artifacts (Borges-Merjane et al., 2020). Another study noted that dissection led to ultrastructural artifacts within 100 µm from the cutting site (Siksou et al., 2007). Handling artifacts that occur during preparation for cryofixation have also been reported to cause detachment of myelin lamellae or separation of axons from the myelin sheath (Möbius et al., 2010). Summary The major limitation of cryofixation methods is their inability to preserve large tissue volumes without ice damage in the sample core. Although regions close to the surface of a tissue sample can be exquisitely preserved in a vitrified state, deeper regions suffer from progressively more severe freezing artifacts due to slower cooling rates. Increasing the pressure during freezing or using cryoprotectants can reduce the critical cooling rate required for vitrification and thereby increase the vitrification depth. The thinness of the samples required for cryofixation is also a challenge for long-term brain banking, as only small pieces of tissue can be stored. Therefore, while cryofixation enables optimal ultrastructural preservation of brain tissue, it is currently limited to relatively small samples, precluding its widespread use in neuroscience research requiring whole brain or thick tissue preservation. Freezing without cryoprotectants Preservation process In the absence of exogenous cryoprotectants or sufficient cooling rates for cryofixation, ice formation during freezing of brain tissue is inevitable. The critical question becomes how much damage to the tissue morphology will occur because of this ice formation. Slower cooling rates allow water to leave cells during the freezing process, causing ice to form predominantly in the extracellular space. Some studies have suggested that more rapid cooling rates are preferred when the goal is to preserve tissue morphology, particularly in specimens with complex microanatomical structures like brain tissue. This is because extracellular ice, while less damaging to cell viability because it helps avoid intracellular ice, can severely disrupt tissue structures, such as cellular processes and synaptic connections (Pegg, 2007). Rapid cooling minimizes the time available for ice crystal nucleation, growth, and rearrangement, which may cause less damage to cellular morphology (Rosene et al., 1986). Therefore, in the context of freezing brain tissue without cryoprotectants, the most common approach used by investigators with the goal of morphological preservation has been to maximize the cooling rate. The main challenge lies in achieving sufficiently rapid cooling throughout the entire tissue volume, particularly for larger specimens. Effects on tissue morphology There were numerous methods described for freezing brain and other nervous tissue in the studies we identified. These included (a) placing the specimen in a freezer, such as one set to ‑80 °C or ‑20 °C (Vonsattel et al., 1995; Kagan and Viner, 2022), (b) placing the tissue in or between a form of dry ice at ‑78.5 °C (Beach et al., 2008), (c) immersing the specimen in liquid nitrogen (Ulfig et al., 1998), (d) immersing the specimen in liquid nitrogen vapor (Davis et al., 2019), (e) immersing the specimen in isopentane that has been pre-cooled with a different cryogen, such as liquid nitrogen or dry ice (Priemer et al., 2023), or (f) freezing the specimen against metal surfaces that have been pre-cooled with a different method, such as liquid nitrogen vapor or a ‑80 °C freezer (Vonsattel et al., 2008). The efficacy of these freezing methods varied considerably in the studies we identified (Table 3; Supplementary File 4) (Rosene and Rhodes, 1990). One study found that freezing human brain slices either with dry ice snow, isopentane pre-cooled in liquid nitrogen, copper plates pre-cooled to ‑80 °C, or placing it in a ‑20 °C freezer after embedding the tissue in a cryo-conductive mixture of chemicals called Optimal Cutting Temperature (OCT) all provided good results, with well-preserved brain tissue architecture and cell morphology as seen on light microscopy. In contrast, samples directly frozen in a ‑80 °C freezer or immersed in liquid nitrogen showed high levels of ice crystal artifacts (Meyronet et al., 2015). A particularly important clinical application of freezing methods is the use of frozen sections. One study found that snap freezing biopsy samples in dry ice-cooled isopentane led to considerably better histology preservation and less ice formation compared to freezing samples by placing them on the freezing shelf within the cryostat at a temperature of around ‑20 °C (Priemer et al., 2023).
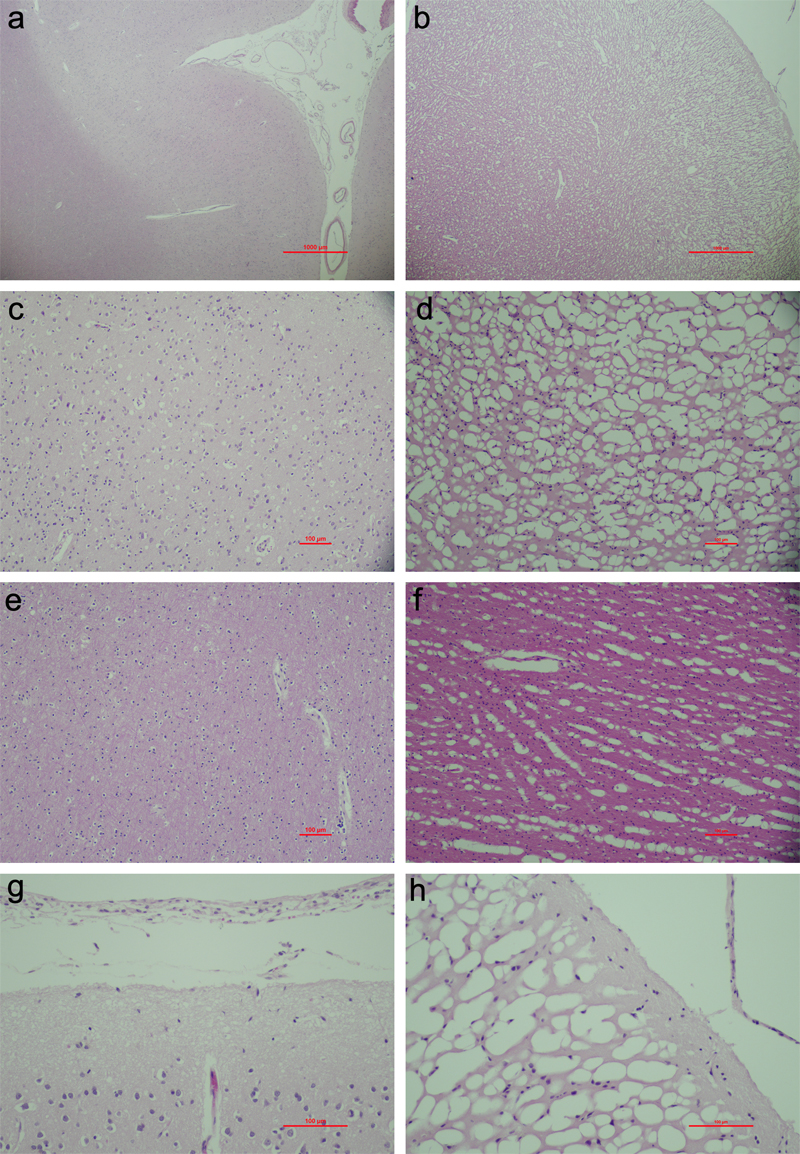
Comparison of methods The differences in histologic outcomes between these freezing methods is thought to largely stem from their different cooling rates, which in turn is related to the properties of the cooling media. Freezer-based methods, while practical for long-term storage, typically offer slow cooling rates because they rely on convective heat transfer via the air, which is a poor thermal conductor. As a result, the cooling process is gradual and less efficient. Immersion of brain slices in liquid nitrogen vapor, although also a gas, has been reported to result in freezing with good morphological preservation (Vonsattel et al., 1995; Davis et al., 2019). This is likely in part because liquid nitrogen vapor is much colder (often between ‑150 °C and ‑196 °C) than a typical ultracold freezer (‑80 °C). The greater temperature differential between the liquid nitrogen vapor and the tissue allows for more rapid heat transfer and cooling, even though nitrogen vapor is still a relatively poor thermal conductor compared to liquids. Dry ice methods provide faster cooling than the use of freezers, but the cooling rate can be inconsistent depending on how well the tissue contacts the dry ice. Otherwise, these methods face the same issue of inefficient heat transfer through gas. Freezing on pre-cooled metal surfaces can achieve very high cooling rates at and close to the freezing surface. One study measured the efficacy of aluminum sheets pre-cooled with dry ice in cooling 1 cm thick fresh brain slices (Nochlin et al., 1993). Using a thermocouple probe, the authors found that a core temperature in the slices of ‑70 °C was reached within 10 minutes. They found no ice crystals in most light micrographs. This result suggests that a sufficiently fast cooling rate was achieved, so that only relatively small ice crystals were formed. However, the rate of freezing when using cooled surfaces is expected to decrease with depth. As a result, methods relying on physical contact with cooled surfaces are more effective for preserving the morphology of surface structures, such as brain slices, but will be less effective for larger specimens, especially if the tissue is not flat. When comparing isopentane and liquid nitrogen immersion, isopentane is generally preferred (Wieser et al., 2022). The key reason is that liquid nitrogen has an extremely low boiling point of ‑196 °C. This means that even a slight degree of warming due to the specimen being plunged into it causes a rapid vaporization that leads to an insulating layer of vapor around the specimen that impedes effective heat transfer out of the specimen. This is known as the Leidenfrost effect. On the other hand, isopentane cooled by a different cryogen remains liquid at temperatures well below its boiling point of 28 °C. This large gap between its cooled temperature and boiling point allows isopentane to maintain direct contact with the specimen without forming a vapor barrier, thus enabling more efficient heat transfer and rapid cooling. In other words, cooled isopentane is not susceptible to the Leidenfrost effect. For this reason, pre-cooled isopentane allows for better thermal contact as well as the potential for very low temperature, making it a commonly preferred option in biobanking. One study measured the rate of cooling using dry ice and dry ice-cooled isopentane in a albumin-gelatin block meant to simulate brain tissue (Rosene et al., 1986). They found that dry ice led to cooling rates of ‑1.5 to ‑1.7 °C/min, while dry-ice cooled isopentane led to cooling rates of ‑3.2 to ‑3.6 °C/min, consistent with isopentane acting as a very effective cryo-conductor. One reported downside of immersion in both isopentane and liquid nitrogen is the higher rate of fractures in brain tissue, as compared to freezing in liquid nitrogen vapor (Vonsattel et al., 1995). As previously discussed, the fractures may be caused by mechanical stress due to sudden temperature changes (Adam et al., 1990). Pulverized dry ice has been reported to be able to cryopreserve brain hemispheres without causing deep crevasses due to opposed temperature fronts in the specimen (Annese, 2004). Damage during tissue rewarming The rewarming phase can significantly impact the final tissue morphology and is often thought to cause more damage than the initial freezing. This is related to the physics of ice formation. At higher subzero temperatures, the probability of forming new ice nuclei is relatively low, while the rate of ice growth is relatively high (Fahy and Wowk, 2015). On the other hand, at low subzero temperatures that are still above the glass transition temperature, the probability of forming new ice nuclei is relatively high, while the rate of ice growth is relatively low. As a result, as the temperature increases during tissue warming, smaller ice nuclei have the opportunity to recrystallize into larger ice crystals (Fahy and Wowk, 2015). Larger ice crystals are expected to cause morphological structural damage. To mitigate ice damage during rewarming, several approaches have been developed. Some sources have reported that more rapid thawing of tissue leads to better outcomes. For example, one study found better neural transplantation outcomes after thawing in a hot water bath at 37 °C for 30–40 seconds, compared with thawing by leaving the neural tissue at room temperature for 5–7 minutes (Das et al., 1983). Another source found high-quality ultrastructural preservation of several non-brain tissue types when blocks were frozen in liquid nitrogen, sectioned into around 5 mm3 blocks on dry ice, and then rapidly thawed at 37 °C in fixative (Galhuber et al., 2021). Alternatively, one protocol involved transferring frozen tissue directly into chilled fixative solutions (Ramirez et al., 2018). This study reported minimal freezing artifacts and well-preserved cell morphology when transferring frozen brain tissue to 1 °C chilled formalin for 72 hours before further processing. The approach of breaking frozen tissue into pieces and immersing it into fixative while the fragments are still frozen, prior to fixation at refrigerator temperature, has also been reported to be effective in rewarming frozen pancreatic tissue (Fortunato et al., 2016). As an additional complication, rewarming frozen tissue may actually decrease the observed burden of histologic alterations in some circumstances. For example, one study using slow freezing of rat cutaneous nerves at ‑10 °C to ‑15 °C with subsequent freeze substitution followed by electron microscopy found that the tissue showed severe ice damage and ultrastructural artifacts (Menz, 1971). Only some ultrastructural features could be recognized, which was attributed to severe dehydration of other structures such as neurofilaments and mitochondria. However, when the slowly frozen tissue was rewarmed via rapid thawing to 40 °C prior to fixation, electron microscopy showed an improved preservation of neurofilaments and mitochondria. Notably, there were still numerous artifacts; for example, the myelin layers were disturbed, and the mitochondria were empty and lacked clearly defined membranes. Mechanistically, the dehydration that prevented the visualization of structures in the freeze-substituted state may have been reversed by thawing. A separate study examined electron microscopy in 1 cm thick sections of human brain tissue that were frozen between dry ice-cooled aluminum plates and immersion-fixed during thawing (Nochlin et al., 1993). The study found that the preservation quality in most images was similar to that in non-frozen brain tissue, with the different cell types and processes easily able to be identified. This finding contradicts the literature using cryofixation and freeze substitution followed by electron microscopy, which, as discussed above, generally finds severe histologic damage in regions deep enough to be affected by ice crystals. One source discusses ultrastructural preservation quality after the cryopreservation of several different cell types, especially hepatic cells, but also with some discussion of brain cells (Trump et al., 1965). The authors note that slow freezing causes significant extracellular ice damage, but that there is a significant amount of what they call “structural restitution” on thawing when cells are preserved with slow freezing. They primarily discuss intracellular features such as mitochondria, which are more likely to be protected by the predominantly extracellular ice formation that will occur during slow freezing. Also, the extent to which any such structural restitution yields the same original morphology present prior to freezing is not entirely clear. Taken together, the issue of histologic damage during rewarming, and how to best decrease or prevent it, appears to be an unresolved one. There is a need for more research comparing different cooling and rewarming methods and for the development of more objective histologic metrics to compare across methods and studies. Tissue freezing in brain banking Perhaps the simplest way of freezing tissue in brain banking is to isolate the whole brain or a brain hemisphere and place it in a freezer. This method was used early in the history of brain banking (Bird, 1980). However, this method has been found to lead to severe ice artifacts on histology, consistent with the slow cooling and rewarming rates expected when freezing and thawing a large brain specimen in this manner (Tourtellotte et al., 1993; Vonsattel et al., 1995, 2008). Cryopreserving very large samples is also impractical for most research applications, because accessing small parts of the brain for study would require several freeze-thaw cycles, leading to even more severe morphological damage. Instead, what appears to be the most common method in modern brain banking is to section fresh brain tissue and use pre-cooled metal plates to freeze the slices (Vonsattel et al., 2008). To assess the degree of morphological preservation in a routinely banked brain preserved via freezing in such a manner, we examined images from cortical tissue that was initially preserved by freezing between metal plates pre-cooled in a ‑80 °C freezer and then rewarmed in cold fixative. We found that in this sample, ice artifacts are readily apparent in the frozen tissue compared to the immersion-fixed tissue (Figure 2). The cells, especially those in the grey matter, are shrunken and mechanically squashed into the pockets of non-frozen tissue between large areas of empty space indicative of ice formation. In this example, the degree of ice artifact in white matter appears to be relatively less severe than in the grey matter. Notably, myelin has previously been reported to be relatively resistant to freezing damage (Möbius et al., 2010).
Figure 2. Example images showing ice artifacts in frozen-thawed brain tissue. Summary Freezing methods are the most common approach for cryopreserving relatively large brain tissue samples. The efficacy of freezing methods varies considerably based on the specific technique used. Placing large tissue samples directly in a freezer leads in general to severe ice artifacts due to the relatively slow cooling rates. On the other hand, freezing brain slices using either pre-cooled metal plates, embedding in dry ice, immersing in cooled isopentane, or freezing in liquid nitrogen vapor, have all been reported to achieve relatively rapid cooling and to maintain morphology with relatively few ice artifacts detectable by light microscopy. Cryoprotectant-based cryopreservation Preservation process Cryoprotectants are substances that can be added to tissues to prevent ice formation during the cryopreservation process. The most frequently used cryoprotectants reported in the studies we identified include dimethyl sulfoxide (DMSO), glycerol, sucrose, dextran, bovine serum albumin, and polyvinylpyrrolidone (PVP). These substances act by forming hydrogen bonds with water molecules, causing freezing point depression, and increasing tissue viscosity. To achieve complete vitrification and prevent ice formation entirely, a high concentration of cryoprotectants is needed. For example, a solution containing 61 % w/v ethylene glycol has been used to vitrify mouse brain slices alongside cooling in liquid nitrogen (German and Akdaş, 2024). Lower concentrations of cryoprotectant have been used in various studies but are not likely to achieve complete vitrification. Notably, cryoprotectants can also cause structural damage. For example, DMSO and glycerol have both been reported to cause ultrastructural alterations to the cell membrane, with DMSO being more damaging than glycerol (McIntyre et al., 1974). Furthermore, cell organelles can be disrupted and myelin-like figures can occur within the cytoplasm. Cryoprotectant immersion Several studies that we identified reported immersion of cryoprotectants into brain tissue prior to cryopreservation (Table 4; Supplementary File 4). Among these, multiple studies found that areas with less take-up of cryoprotectant tend to have more freezing artifacts, but that in areas where an adequate concentration of cryoprotectant was successfully taken up, cells are undamaged (Seilhean et al., 1996; Pichugin et al., 2006). Another study demonstrated a potential issue with cryoprotectant immersion, as cellular necrosis occurred during the time necessary for the immersion procedure to be completed (German and Akdaş, 2024).
Cryoprotectant perfusion Perfusion is an alternative method for delivering cryoprotectants through the cerebrovascular system that can potentially achieve more uniform distribution. Several studies that we identified used perfusion of cryoprotectants through the blood vessels of the brain in laboratory animals (Table 5; Supplementary File 4). One study tested perfusion of rat brains with either a vitrification solution called VM1 or a solution containing 13 % DMSO and 13 % glycerol (Canatelli-Mallat et al., 2020). While no ice formation was observed in either group on light microscopy, significant shrinkage of neurons occurred. Another study used perfusion of mouse brains with 30 % sucrose for 15 minutes, followed by overnight immersion in 30 % sucrose under refrigeration before freezing in isopentane/2-methylbutane cooled with liquid nitrogen (Comba et al., 2020). This protocol was found to preserve morphology in tumor tissue within the brain as seen on light microscopy, with no histologically detectable ice crystals. The extended immersion period makes sense because perfusion of the brain is not completely reliable, even in ideal, controlled laboratory conditions (Cahill et al., 2012). Although the latter procedure also raises concerns about potential tissue decomposition, the authors did not report that the extended immersion period affected tissue morphology, consistent with previous literature suggesting that the cellular effects of postmortem decomposition have a relatively slow onset, especially in refrigerated tissue (Krassner et al., 2023). However, cryoprotectant perfusion is expected to be more challenging in a brain banking context, largely as a result of agonal and postmortem changes (McFadden et al., 2019; McKenzie et al., 2022).
Summary Cryoprotectants like DMSO, glycerol, and sucrose can reduce ice formation during cryopreservation and potentially enable vitrification. High cryoprotectant concentrations are needed for complete vitrification, which can cause osmotic stress if the procedure is not performed in a properly graded manner. Immersion of brain tissue in cryoprotectants has been reported to preserve ultrastructure, but certain areas can still have freezing artifacts if cryoprotectant diffusion or uptake by cells are incomplete. Perfusion of cryoprotectants through the cerebrovascular system may enable more uniform cryoprotectant distribution and better tissue preservation in larger brain samples, but this approach is challenging to perform, especially in postmortem brain banking settings, and the evidence for its efficacy in the published literature is currently limited. Storage temperature and length One previous source has comprehensively reviewed the literature on cryostorage (Hubel et al., 2014). The authors report that unfixed biospecimens kept above the tissue’s glass transition temperature are expected to show degradation, even at the ultra-low freezer temperature of ‑80 °C. There are several reasons for this. First, the presence of residual liquid water allows for ongoing molecular motion. Second, degradative biomolecules, such as proteases, nucleases, and lipases, may still have activity, which can lead to biochemical alterations. Although the activity of degradative enzymes is expected to be greatly reduced, it is not necessarily completely halted at ‑80 °C, and this can add up over long periods of storage. Third, ice crystals will become larger during long-term storage, leading to more damage to tissue morphology (Choi et al., 2015; Zennoune et al., 2022). Such ice recrystallization is expected to be especially common as a result of temperature fluctuations (Petzold and Aguilera, 2009). All these factors add up to potentially damage tissue during storage at temperatures above the glass transition temperature. This has been borne out empirically in some, but not all studies. One study found that human brain tissue stored at ‑70 °C for more than one year developed irregular spaces and variations in section thickness that hindered comparative analyses (Itoyama et al., 1980). Another study of dog brain samples held at ‑18 °C found that this led to increased damage to neurons and to increased large, irregular, clear areas under the microscope after 7 days, as compared to 2 days of storage (Baraibar and Schoning, 1985). Notably, in this study, hemolysis was already complete by 2 days, so it did not worsen by 7 days of storage. However, other studies have found that histomorphological features were not altered after years of storage of brain tissue at ‑80 °C (Zilles et al., 2002; Andreasson et al., 2013). Taken together, these findings underscore the importance of considering storage length as a potential confounding variable when analyzing banked specimens, although whether it makes a large difference to the morphology in practice is still an open question and likely depends on the variable of interest. Storage below the glass transition temperature, on the other hand, is expected to cause fewer changes. For example, one study found that while liver cell spheroids stored at ‑80 °C had a loss of viable cells starting at one month and progressing substantially up until one year, storage in liquid nitrogen vapor at approximately ‑170 °C led to no changes in viability (Massie et al., 2013). Another source using storage at ‑180 °C found that morphology did not vary over the storage period of a few days to up to more than 5 years (Seilhean et al., 1996). However, as previously discussed, storage at this temperature can cause a buildup of fractures (Hunt et al., 1994). Perhaps more importantly, storage at temperatures below ‑80 °C would incur a significant cost that may not be justified for many brain banks. It is also worthwhile to point out that any freeze-thaw cycle is likely to be extremely damaging to tissue morphology. One study found that a freeze-thaw cycle softened brain tissue to such a degree that it was too difficult to process it for histologic study at all (Schäfer and Kaufmann, 1999). The authors noted that this phenomenon was common in tissues like the brain that have a relatively high extracellular water content and relatively weak physical connections between cells. This is one of the reasons that brain banks typically break up fresh tissue into smaller blocks prior to the initial freezing procedure (Shepherd et al., 2019). Relationship between ice crystal formation and postmortem decomposition It is mechanistically plausible that a longer postmortem interval could increase ice artifacts, for two reasons. First, the levels of free water have been found to increase in the brain after death (Ansari et al., 1976; Leonard et al., 2016). The increased water content may make tissue more susceptible to ice crystal formation during the freezing process. Second, the molecular links maintaining tissue structures break down during the postmortem interval (Krassner et al., 2023). This may render tissue structures more vulnerable to the mechanical stresses induced by ice crystal growth. This relationship is also consistent with the findings in frozen sections that tissue edema tends to worsen ice artifacts (Tofte et al., 2014; Priemer et al., 2023). Notably, astrocytes tend to accumulate water in the postmortem interval, and electron microscopy has found that astrocyte membranes are the most vulnerable to freezing damage, which is particularly perceptible around neurons, oligodendrocytes, and capillaries (Walder and Vrensen, 1972). One study found that the method of tissue preservation affected how a 20 hour postmortem delay affects myelin basic protein immunostaining (Itoyama et al., 1980). When using immersion fixation, the authors found that some myelin sheaths became vacuolated after this postmortem delay, but the overall preservation was satisfactory. On the other hand, tissue that was frozen after the same postmortem delay showed more severe changes, including few visible oligodendrocyte processes and only faintly stained myelin sheaths. Although the hypothesis of a relationship between postmortem interval and ice damage is interesting, it has not been widely investigated and it is worthy of more research before too much confidence is placed into these preliminary observations. Another relevant factor is that some structural changes observed in frozen tissue can overlap with those seen in postmortem autolysis. Mechanistically, both freezing and postmortem decomposition would be expected to cause cell membrane damage. For example, one study noted that hemolysis, characterized by the loss of erythrocyte cytoplasm, was present in both frozen-thawed and decomposed fish brain tissue (Kagan and Viner, 2022). Another study, comparing the ultrastructural alterations in larval fly brains exposed to either lethal freezing or anoxia, found similarities such as nuclear membrane expansions, nuclear chromatin clumping, and rough endoplasmic reticulum swelling (Lee et al., 1997). These findings suggest that some of the damage observed in frozen brain tissue may be exacerbated by – or difficult to distinguish from – the changes that occur during the postmortem interval. Effects of cryopreservation on biomolecular preservation Modern brain banking aims to preserve multiple tissue properties, and many tissue requests are for molecular analyses, making this aspect of preservation quality also critical to consider. From a mechanistic perspective, cryopreservation is not thought to cause the formation or breakdown of covalent bonds in biomolecules (Fahy and Wowk, 2015), which is an obvious advantage over chemical fixation by aldehydes for the purpose of biomolecular assays. However, cryopreservation can cause numerous other effects on biomolecules, including phase changes in membrane lipids, conformational changes due to loss of the hydration shell, hydrophobic aggregation of biomolecules, and protein inactivation, for example of the sodium–potassium pump (Jain et al., 2021; Pegg, 1976). Some biomolecular changes occur during the cryopreservation process because metabolism does not fully stop until very low temperatures are reached. For this reason, faster cooling processes might be expected to yield superior biomolecular preservation. Indeed, for tumor tissues, snap freezing of tumor biopsies has been reported to be the gold standard for maintenance of biomolecular properties of tissues, such as their content of nucleic acids and phosphoproteins (van der Wijngaart et al., 2023). However, other data have suggested that slow freezing may be preferable for maintaining biomolecular properties, because fast freezing yields small ice crystals with an increased contact area between ice and liquid, which may increase the net exposure of proteins to the damaging effects of ice (Cao et al., 2003). One potential solution to the problem of biomolecular damage due to ice formation would be to use cryoprotectants, since a sufficient concentration of cryoprotectants could prevent ice damage. Cryoprotectants could also allow for better maintenance of cellular integrity, which could be useful for the molecular profiling of single cells. However, if cryoprotectants are used, they also have the potential to affect the molecular properties of the tissue itself, for instance by causing protein denaturation (Fahy and Wowk, 2015). For some molecular assays, cryoprotectants used at potentially variable concentrations within or between samples could also create batch effects that would confound subsequent analyses. In practical brain banking settings, cryopreservation of small tissue blocks in isopentane cooled by liquid nitrogen has been reported to be a preferred method for obtaining high-quality RNA and protein preservation (Durrenberger et al., 2010). However, different freezing methods have been found not to have a significant effect on the quality of RNA harvested from brain tissue (Durrenberger et al., 2010). Additionally, one study found that there was no difference between three different freezing methods in the protein profiles detected by mass spectrometry (Meyronet et al., 2015), even though there were differences in the degrees of morphological preservation. It seems that differences in cryopreservation protocols have smaller effects on many biomolecular features than on histology. For nucleic acids in particular, studies have found that proxies of a prolonged and severe agonal state, such as hospitalization prior to death or decreased brain pH, are the strongest correlate of worsened RNA quality (Li et al., 2004, 2007; Durrenberger et al., 2010; Hagenauer et al., 2018). Comparison to other tissues Several studies have noted that the brain appears to be more vulnerable to freezing damage than other organs and tissues. One source found that while kidney tissue did not show substantial changes after freezing, brain tissue softened significantly, making it difficult to process the brain tissue for evaluation (Schäfer and Kaufmann, 1999). These authors hypothesized that tissue softening after freezing occurs in tissues with high extracellular water content and with relatively weak physical connections holding the tissue together. They also noted that during slow freezing, extracellular ice formation can cause cells to shrink significantly, potentially tearing apart cell-cell connections, which will not redevelop upon thawing. Another source observed that freezing damage in the brain seemed to be worse compared to other tissues (Lee et al., 1997). The authors noted that the brain appears to be the “weak link” with the most alterations following freezing in the fly larvae they were studying. Finally, one cryofixation study noted that nervous system tissue is more difficult to vitrify than any other tissue they had studied (Zuber et al., 2005). The authors reported that while tissues like cartilage or skin could be vitrified without cryoprotectants, this was not possible for nervous tissue, which required the use of cryoprotectants for successful vitrification via cryofixation. The authors attributed this difficulty to the brain's high content of water and lower intrinsic effect of cryoprotectants. Collectively, these findings suggest that the unique composition and structure of brain tissue makes it particularly challenging to cryopreserve brain tissue effectively as compared to many other tissue types. Comparison to previous reviews To the best of our knowledge, there has not been a previous comprehensive review primarily on the cryopreservation of brain tissue. There have been many previous reviews on cryobiology in general, several of which can be highlighted (Bojic et al., 2021; Fahy and Wowk, 2021; Murray and Gibson, 2022). In recent years there has also been an increased focus on cryopreserving complex tissues or organs (Giwa et al., 2017). However, these reviews have not focused on cryopreservation of brain tissue, which is not currently considered to be as important as other tissue types for clinical applications such as transplantation. The reviews on brain tissue cryopreservation have tended to focus on aspects other than histology, such as biobanking in general (Shabihkhani et al., 2014) or cellular survival (Paynter, 2008). Notably, one review notes that most techniques on nerve cell cryopreservation have been derived from those applied to cells of other organs, rather than being optimized for nerve cells in particular (Paynter, 2008). One review article primarily focusing on biochemical and biomolecular studies in banked brain tissue also reports that frozen brain tissue can been used in detailed morphological studies (Hynd et al., 2003). Finally, one review on the practice of cryonics discusses the cryopreservation of the brain (Best, 2008). The author primarily discusses articles describing successful preservation of brain tissue when cryoprotectants are used. Compared to our review, the aforementioned review does not extensively discuss studies on the histological outcomes of freezing without cryoprotectants, which will occur if the cryoprotectant solution does not reach all areas of the brain. Limitations of this review This review has a number of limitations that should be considered. First, the data reported in some articles were occasionally ambiguous, making it difficult to tell whether the observations were based on first-hand experience or on second-hand reports. While we attempted to include only primary data, this ambiguity may have introduced some second-hand data into our findings. Second, our review focused primarily on brain tissue cryopreservation, with limited consideration of other tissue types. Given the potential similarities across tissues, a more comprehensive review incorporating the broader literature could provide additional insights. Third, the “grey” literature on this technical topic, such as information shared on mailing lists and platforms like ResearchGate, was not included in our review. Future reviews may benefit from carefully considering these unpublished data, as they may contain valuable insights from researchers who have addressed the topic in practice without formally publishing their findings. Fourth, the distinctions between the different categories of cryopreservation techniques of cryofixation, freezing, and cryopreservation with cryoprotectants can be somewhat blurred, as some studies may employ methods that overlap or share similarities across these categories. For example, some cryofixation studies use cryoprotectants. This potential ambiguity in categorization may have influenced the interpretation and synthesis of the reviewed studies. Fifth, our assessment of preservation quality relies heavily on the verbal reports and descriptions provided by the authors of the reviewed studies. The lack of standardized, quantitative metrics for evaluating preservation quality across studies limits the objectivity and comparability of these assessments. Overall, the literature on morphological outcomes following cryopreservation of unfixed brain tissue is relatively limited compared to studies using fixed tissue, particularly outside of the cryofixation domain. The majority of the reviewed studies have small sample sizes and limited replication, which may affect the generalizability of their findings. As a result, the conclusions drawn in this review should be interpreted with caution, as they are based on a relatively small number of studies. Further research with larger sample sizes and standardized evaluation methods is needed to corroborate and extend the findings of this review. Summary of cryopreservation methods discussed We can summarize several of the cryopreservation methods discussed in this article (Table 6). For smaller specimens such as tissue biopsy samples, cryofixation or cryoprotectant-based methods likely offer the best histologic preservation, especially as viewed under the electron microscope. However, for the larger whole brain or hemisphere slices that are typically preserved in brain banking, freezing methods are the only published methods that have been reported to be reliably effective. Among the cryopreservation methods for brain slices, freezing between cooled metal plates, embedding in dry ice, or immersion in cooled isopentane appear to be the most effective for preserving relatively large samples while minimizing ice artifacts as seen on light microscopy (Vonsattel et al., 2008; Meyronet et al., 2015). Notably, many investigators do not focus on detailed morphological preservation for their frozen tissue, focusing instead on preservation of biochemical properties, making the histologic considerations less important.
Implications for brain banking Brain banks may have different needs for their cryopreservation methods depending upon the intended use of their tissue samples (Table 7). For whole brain slices, which are commonly used in brain banking, preservation of overall tissue architecture is often a primary concern. In this case, freezing the slices between cooled metal plates offers a relatively simple and effective approach. For smaller tissue fragments, where preservation of cell membrane morphology as seen on light microscopy is desired, rapid freezing methods such as immersion in liquid nitrogen vapor or liquid nitrogen-cooled isopentane can be used. However, the effectiveness of these methods for preserving delicate cellular structures can vary depending on factors such as tissue type and freezing rate. If one has access to specialized equipment and expertise, then cryofixation could potentially be used on very thin sections from small biopsy specimens to achieve high-quality preservation of subcellular structures such as synaptic vesicles. For those more interested in preserving the biomolecular properties of the tissue and not as much in detailed histological preservation, the choice of freezing method is less critical. In this case, simpler methods, such as placement in dry ice or in an ultra-low temperature (‑80 °C) freezer, might be sufficient. Finally, for specialized research applications, such as those interested in the preservation of electrophysiological features of fresh tissue slices (German and Akdaş, 2024), the use of cryoprotective agents, while a relatively complex procedure, might be justified. Because cryopreservation can be expensive and highly labor-intensive, it is essential to be pragmatic and to choose the simplest method that adequately meets the specific preservation needs of the brain bank and its research community. This approach ensures efficient use of resources while still maintaining the quality necessary for intended downstream applications.
We can also consider the perspective of researchers using existing tissue banked at brain banks. Samples that are frozen may have more variable histologic qualities than commonly appreciated. For researchers interested in histological features, it may be worthwhile to seek tissue that has been frozen in a rapid, well-controlled manner, rather than assuming that the morphological quality of all frozen tissue will be identical. Years of storage at ‑80 °C also has the potential to affect tissue quality. Using rapid thawing techniques and/or thawing in fixative may facilitate morphological preservation in research applications, although the best methods for tissue rewarming are still not well established. Notably, preservation of histologic features is generally expected to be superior in banked tissue that has been chemically fixed. Therefore, there is currently a trade-off between the types of tissue generally available from brain banks, with frozen samples allowing certain molecular biology studies to be more easily performed, and fixed samples expected to have better morphological preservation. Areas for further research Some research applications, such as long-distance circuit mapping and also biochemical assays, could potentially benefit from the ability to cryopreserve an entire brain while maintaining its structural integrity. However, current methods struggle with cryopreserving large tissue volumes without ice damage. Future studies could explore improved methods for cryoprotectant perfusion of the brain, which has received relatively little attention in the published literature over the years (Suda et al., 1966; Lemler et al., 2004; Canatelli-Mallat et al., 2020; Comba et al., 2020). Corroborating and potentially improving such efforts could provide insights into the feasibility and the limitations of whole brain cryopreservation via cryoprotectant perfusion. Several open questions also remain regarding the basic mechanisms and limits of ice damage during cryopreservation. Research is needed to better characterize the ice damage caused both during the initial cooling of brain tissue and during the rewarming of large brain samples. Technological approaches to increase the speed of rewarming, such as dielectric warming (Wowk et al., 2024) or ultrasound rewarming (Alcalá et al., 2023), may have the potential to decrease damage in cryopreserved brain tissue further, although this requires more study. Finally, the use of computational methods to infer the most probable original state of ice-damaged tissue is an area for future work. Some progress has already been made in this direction for frozen sections (Ozyoruk et al., 2022). However, in this study, the algorithm could not reverse the alterations of cellular positions when severe ice artifacts were present. There is a need for a better theoretical understanding of the limits of such computational inference methods to reverse ice damage. Conclusions This review highlights the complex landscape of brain tissue cryopreservation, revealing both the potential and limitations of current methods. Key themes that emerged from our literature review included the relationship between cooling rate and freezing injury, the effects of different cryoprotectants, and the impact of tissue characteristics on cryopreservation outcomes. Of the study types identified, cryofixation offers exceptional ultrastructural preservation but is limited to very thin samples, while freezing approaches can achieve minimal ice artifacts as seen on light microscopy in relatively larger specimens, particularly when rapid cooling methods are employed. The use of cryoprotectants shows promise for high-quality preservation, but remains challenging for achieving consistently even distribution, especially in larger samples and postmortem settings. Notably, brain tissue appears to be particularly vulnerable to freezing damage as compared to other tissues, underscoring the need for more research in this area. While current cryopreservation techniques are sufficient for many research applications, there remains an unmet need for methods capable of preserving larger brain samples with minimal structural disruption. Future research may benefit from a focus on improving cryoprotectant delivery, optimizing cooling and rewarming protocols, and exploring computational methods to infer the original state of the tissue despite the presence of freezing artifacts. Abbreviations DMSO - Dimethyl sulfoxide; OCT - Optimal Cutting Temperature compound; PMI - Postmortem interval; TEM - Transmission electron microscopy; VM1 and VM3 - Vitrification solutions composed of multiple cryoprotectants. Acknowledgements We would like to thank Brian Wowk and Kenneth Hayworth for helpful personal communications regarding this topic. Data availability This article was primarily built upon publicly available data. The relevant data and analyses are presented in the article or in the supplementary materials. Author contributions A.T.M., E.F.K., K.F., and J.C. conceptualized the article. A.T.M. performed article searching and screening. A.T.M., O.N., and B.W. performed data extraction and review of data extraction. E.T. performed experimental studies and imaging of frozen-thawed brain tissue. A.T.M. wrote the initial draft of the manuscript. All authors reviewed the manuscript. All authors approved the final manuscript. Conflicts of interest statement A.T.M. is an employee of Oregon Brain Preservation, a non-profit brain preservation organization, and a director of Apex Neuroscience, a non-profit research organization. B.W. is affiliated with the European Institute for Brain Research, a non-profit research foundation, and the Biopreservation Institute, a non-profit research organization that works together with Nectome Inc., which plans to offer aldehyde-based biostasis. E.F.K. is a shareholder and CEO of Tomorrow Biostasis, a biostasis provider, and President of the Board of the European Biostasis Foundation, a non-profit research foundation. Funding statement Borys Wróbel’s research is supported by a grant from the Biomedical Research and Longevity Society. Otherwise, the authors received no specific funding for this work. Supplementary Files Supplementary File 1 [download] - RAMESES Checklist [pdf format]. Supplementary File 2 [download] - Abstract screening results [csv format]. Supplementary File 3 [download] - Full text screening results [csv format]. Supplementary File 4 [download] - Extracted data table [xlsx format]. References Adam, M., Hu, J.F., Lange, P., Wolfinbarger, L., 1990. The effect of liquid nitrogen submersion on cryopreserved human heart valves. Cryobiology 27, 605–614. https://doi.org/10.1016/s0011-2240(05)80028-8 Alcalá, E., Encabo, L., Barroso, F., Puentes, A., Risco, I., Risco, R., 2023. Sound waves for solving the problem of recrystallization in cryopreservation. Sci. Rep. 13, 7603. https://doi.org/10.1038/s41598-023-34681-z Andreasson, A., Kiss, N.B., Juhlin, C.C., Höög, A., 2013. Long-term storage of endocrine tissues at -80 °C does not adversely affect RNA quality or overall histomorphology. Biopreservation Biobanking 11, 366–370. https://doi.org/10.1089/bio.2013.0038 Annese, J., 2004. Neo-architectonics: Computer aided histological examination of the human visual cortex (Ph.D.). Dartmouth College, United States -- New Hampshire. Ansari, K.A., Rand, A., Hendrickson, H., Bentley, M.D., 1976. Qualitative and quantitative studies on human myelin basic protein in situ with respect to time interval between death and autopsy. J. Neuropathol. Exp. Neurol. 35, 180–90. https://doi.org/10.1097/00005072-197603000-00005 Baraibar, M.A., Schoning, P., 1985. Effects of freezing and frozen storage on histological characteristics of canine tissues. J. Forensic Sci. 30, 439–447. Beach, T.G., Sue, L.I., Walker, D.G., Roher, A.E., Lue, L., Vedders, L., Connor, D.J., Sabbagh, M.N., Rogers, J., 2008. The Sun Health Research Institute Brain Donation Program: Description and Eexperience, 1987–2007. Cell Tissue Bank. 9, 229–245. https://doi.org/10.1007/s10561-008-9067-2 Benson, T.E., Burd, G.D., Greer, C.A., Landis, D.M., Shepherd, G.M., 1985. High-resolution 2-deoxyglucose autoradiography in quick-frozen slabs of neonatal rat olfactory bulb. Brain Res. 339, 67–78. https://doi.org/10.1016/0006-8993(85)90622-5 Best, B.P., 2008. Scientific justification of cryonics practice. Rejuvenation Res. 11, 493–503. https://doi.org/10.1089/rej.2008.0661 Bird, E., 1980. A brain tissue resource center to promote research in schizophrenia. Perspect. Schizophr. Res. Raven N. Y. Bojic, S., Murray, A., Bentley, B.L., Spindler, R., Pawlik, P., Cordeiro, J.L., Bauer, R., de Magalhães, J.P., 2021. Winter is coming: the future of cryopreservation. BMC Biol. 19, 56. https://doi.org/10.1186/s12915-021-00976-8 Borges-Merjane, C., Kim, O., Jonas, P., 2020. Functional Electron Microscopy, “Flash and Freeze,” of Identified Cortical Synapses in Acute Brain Slices. Neuron 105, 992-1006.e6. https://doi.org/10.1016/j.neuron.2019.12.022 Cahill, L.S., Laliberté, C.L., Ellegood, J., Spring, S., Gleave, J.A., Eede, M.C. van, Lerch, J.P., Henkelman, R.M., 2012. Preparation of fixed mouse brains for MRI. NeuroImage 60, 933–939. https://doi.org/10.1016/j.neuroimage.2012.01.100 Canatelli-Mallat, M., Lascaray, F., Entraigues-Abramson, M., Portiansky, E.L., Blamaceda, N., Morel, G.R., Goya, R.G., 2020. Cryopreservation of a Human Brain and Its Experimental Correlate in Rats. Rejuvenation Res. 23, 516–525. https://doi.org/10.1089/rej.2019.2245 Cao, E., Chen, Y., Cui, Z., Foster, P.R., 2003. Effect of freezing and thawing rates on denaturation of proteins in aqueous solutions. Biotechnol. Bioeng. 82, 684–690. https://doi.org/10.1002/bit.10612 Chambers, R., Hale, H.P., Hardy, W.B., 1932. The formation of ice in protoplasm. Proc. R. Soc. Lond. Ser. B Contain. Pap. Biol. Character 110, 336–352. https://doi.org/10.1098/rspb.1932.0029 Choi, Y.-S., Ku, S.-K., Jeong, J.-Y., Jeon, K.-H., Kim, Y.-B., 2015. Changes in Ultrastructure and Sensory Characteristics on Electro-magnetic and Air Blast Freezing of Beef during Frozen Storage. Korean J. Food Sci. Anim. Resour. 35, 27–34. https://doi.org/10.5851/kosfa.2015.35.1.27 Clarke, A., Morris, G.J., Fonseca, F., Murray, B.J., Acton, E., Price, H.C., 2013. A Low Temperature Limit for Life on Earth. PloS One 8, e66207. https://doi.org/10.1371/journal.pone.0066207 Comba, A., Dunn, P.J., Kish, P.E., Kadiyala, P., Kahana, A., Castro, M.G., Lowenstein, P.R., 2020. Laser Capture Microdissection of Glioma Subregions for Spatial and Molecular Characterization of Intratumoral Heterogeneity, Oncostreams, and Invasion. J. Vis. Exp. JoVE. https://doi.org/10.3791/60939 Das, G.D., Houle, J.D., Brasko, J., Das, K.G., 1983. Freezing of neural tissues and their transplantation in the brain of rats: technical details and histological observations. J. Neurosci. Methods 8, 1–15. https://doi.org/10.1016/0165-0270(83)90047-x Davis, S., Scott, C., Ansorge, O., Fischer, R., 2019. Development of a Sensitive, Scalable Method for Spatial, Cell-Type-Resolved Proteomics of the Human Brain. J. Proteome Res. 18, 1787–1795. https://doi.org/10.1021/acs.jproteome.8b00981 Dumoux, M., Glen, T., Smith, J.L.R., Ho, E.M.L., Perdigão, L.M.A., Pennington, A., Klumpe, S., Yee, N.B.Y., Farmer, D.A., Lai, P.Y.A., Bowles, W., Kelley, R., Plitzko, J.M., Wu, L., Basham, M., Clare, D.K., Siebert, C.A., Darrow, M.C., Naismith, J.H., Grange, M., 2023. Cryo-plasma FIB/SEM volume imaging of biological specimens. eLife 12, e83623. https://doi.org/10.7554/eLife.83623 Durrenberger, P.F., Fernando, S., Kashefi, S.N., Ferrer, I., Hauw, J.-J., Seilhean, D., Smith, C., Walker, R., Al-Sarraj, S., Troakes, C., Palkovits, M., Kasztner, M., Huitinga, I., Arzberger, T., Dexter, D.T., Kretzschmar, H., Reynolds, R., 2010. Effects of antemortem and postmortem variables on human brain mRNA quality: a BrainNet Europe study. J. Neuropathol. Exp. Neurol. 69, 70–81. https://doi.org/10.1097/NEN.0b013e3181c7e32f Estrada, L.I., Robinson, A.A., Amaral, A.C., Giannaris, E.L., Heyworth, N.C., Mortazavi, F., Ngwenya, L.B., Roberts, D.E., Cabral, H.J., Killiany, R.J., Rosene, D.L., 2017. Evaluation of long-term cryostorage of brain tissue sections for quantitative histochemistry. J. Histochem. Cytochem. 65, 153–171. https://doi.org/10.1369/0022155416686934 Fahy, G.M., Wowk, B., 2021. Principles of Ice-Free Cryopreservation by Vitrification. Methods Mol. Biol. Clifton NJ 2180, 27–97. https://doi.org/10.1007/978-1-0716-0783-1_2 Fahy, G.M., Wowk, B., 2015. Principles of cryopreservation by vitrification. Methods Mol. Biol. Clifton NJ 1257, 21–82. https://doi.org/10.1007/978-1-4939-2193-5_2 Fortunato, F., Hackert, T., Büchler, M.W., Kroemer, G., 2016. Retrospective electron microscopy: Preservation of fine structure by freezing and aldehyde fixation. Mol. Cell. Oncol. 3, e1251382. https://doi.org/10.1080/23723556.2016.1251382 Galhuber, M., Kupper, N., Dohr, G., Gauster, M., Kwapiszewska, G., Olschewski, A., Jandl, K., Gschwandtner, E., Schweiger, M., Kratky, D., Leitinger, G., Prokesch, A., Kolb, D., 2021. Simple method of thawing cryo-stored samples preserves ultrastructural features in electron microscopy. Histochem. Cell Biol. 155, 593–603. https://doi.org/10.1007/s00418-020-01952-z Gasperoni, J., Dworkin, S., 2024. Cryosectioning and Immunohistochemistry Using Frozen Adult Murine Brain Neural Tissue. Methods Mol. Biol. Clifton NJ 2746, 155–163. https://doi.org/10.1007/978-1-0716-3585-8_13 Gellhaar, S., Sunnemark, D., Eriksson, H., Olson, L., Galter, D., 2017. Myeloperoxidase-immunoreactive cells are significantly increased in brain areas affected by neurodegeneration in Parkinson’s and Alzheimer’s disease. Cell Tissue Res. 369, 445–454. https://doi.org/10.1007/s00441-017-2626-8 Gerber, D., Wilen, L.A., Poydenot, F., Dufresne, E.R., Style, R.W., 2022. Stress accumulation by confined ice in a temperature gradient. Proc. Natl. Acad. Sci. U. S. A. 119, e2200748119. https://doi.org/10.1073/pnas.2200748119 German, A., Akdaş, E.Y., 2024. Excitability and synaptic transmission after vitrification of mouse corticohippocampal slices. https://doi.org/10.1101/2024.06.03.597218 Gilkey, J.C., Staehelin, L.A., 1986. Advances in ultrarapid freezing for the preservation of cellular ultrastructure. J. Electron Microsc. Tech. 3, 177–210. https://doi.org/10.1002/jemt.1060030206 Giwa, S., Lewis, J.K., Alvarez, L., Langer, R., Roth, A.E., Church, G.M., Markmann, J.F., Sachs, D.H., Chandraker, A., Wertheim, J.A., Rothblatt, M., Boyden, E.S., Eidbo, E., Lee, W.P.A., Pomahac, B., Brandacher, G., Weinstock, D.M., Elliott, G., Nelson, D., Acker, J.P., Uygun, K., Schmalz, B., Weegman, B.P., Tocchio, A., Fahy, G.M., Storey, K.B., Rubinsky, B., Bischof, J., Elliott, J.A.W., Woodruff, T.K., Morris, G.J., Demirci, U., Brockbank, K.G.M., Woods, E.J., Ben, R.N., Baust, J.G., Gao, D., Fuller, B., Rabin, Y., Kravitz, D.C., Taylor, M.J., Toner, M., 2017. The promise of organ and tissue preservation to transform medicine. Nat. Biotechnol. 35, 530–542. https://doi.org/10.1038/nbt.3889 Guo, J., Wang, G., Tang, W., Song, D., Wang, X., Hong, J., Zhang, Z., 2020. An optimized approach using cryofixation for high-resolution 3D analysis by FIB-SEM. J. Struct. Biol. 212, 107600. https://doi.org/10.1016/j.jsb.2020.107600 Hagenauer, M.H., Schulmann, A., Li, J.Z., Vawter, M.P., Walsh, D.M., Thompson, R.C., Turner, C.A., Bunney, W.E., Myers, R.M., Barchas, J.D., Schatzberg, A.F., Watson, S.J., Akil, H., 2018. Inference of cell type content from human brain transcriptomic datasets illuminates the effects of age, manner of death, dissection, and psychiatric diagnosis. PLOS ONE 13, e0200003. https://doi.org/10.1371/journal.pone.0200003 Han, B., Bischof, J.C., 2004. Direct cell injury associated with eutectic crystallization during freezing. Cryobiology 48, 8–21. https://doi.org/10.1016/j.cryobiol.2003.11.002 Harreveld, A.V., Fifkova, E., 1975. Rapid freezing of deep cerebral structures for electron microscopy. Anat. Rec. 182, 377–385. https://doi.org/10.1002/ar.1091820311 Heuser, J.E., Reese, T.S., Landis, D.M., 1976. Preservation of synaptic structure by rapid freezing. Cold Spring Harb. Symp. Quant. Biol. 40, 17–24. https://doi.org/10.1101/sqb.1976.040.01.004 Hirokawa, N., Kirino, T., 1980. An ultrastructural study of nerve and glial cells by freeze-substitution. J. Neurocytol. 9, 243–254. https://doi.org/10.1007/BF01205160 Hisano, S., Adachi, T., Maegawa, M., Daikoku, S., 1986. Some improvement in tissue preparation and colloidal-gold immunolabeling for electron microscopy. Am. J. Anat. 175, 245–266. https://doi.org/10.1002/aja.1001750210 Hubel, A., Spindler, R., Skubitz, A.P.N., 2014. Storage of human biospecimens: selection of the optimal storage temperature. Biopreservation Biobanking 12, 165–175. https://doi.org/10.1089/bio.2013.0084 Hunt, C.J., Song, Y.C., Bateson, E.A., Pegg, D.E., 1994. Fractures in cryopreserved arteries. Cryobiology 31, 506–515. https://doi.org/10.1006/cryo.1994.1061 Hynd, M.R., Lewohl, J.M., Scott, H.L., Dodd, P.R., 2003. Biochemical and molecular studies using human autopsy brain tissue. J. Neurochem. 85, 543–562. https://doi.org/10.1046/j.1471-4159.2003.01747.x Itoyama, Y., Sternberger, N.H., Kies, M.W., Cohen, S.R., Richardson, E.P., Webster, H., 1980. Immunocytochemical method to identify myelin basic protein in oligodendroglia and myelin sheaths of the human nervous system. Ann. Neurol. 7, 157–166. https://doi.org/10.1002/ana.410070211 Jain, K., Salamat-Miller, N., Taylor, K., 2021. Freeze-thaw characterization process to minimize aggregation and enable drug product manufacturing of protein based therapeutics. Sci. Rep. 11, 11332. https://doi.org/10.1038/s41598-021-90772-9 Jones, E.G., Hendry, S.H.C., Liu, X.-B., Hodgins, S., Potkin, S.G., Tourtellotte, W.W., 1992. A method for fixation of previously fresh-frozen human adult and fetal brains that preserves histological quality and immunoreactivity. J. Neurosci. Methods 44, 133–144. https://doi.org/10.1016/0165-0270(92)90006-Y Kagan, R., Viner, T.C., 2022. Identification of freeze-thaw artifact in fresh and decomposed black rockfish (Sebastes melanops) and steelhead trout (Oncorhynchus mykiss). Vet. Pathol. 59, 1022–1030. https://doi.org/10.1177/03009858221120012 Karlsson, J.O., Toner, M., 1996. Long-term storage of tissues by cryopreservation: critical issues. Biomaterials 17, 243–256. https://doi.org/10.1016/0142-9612(96)85562-1 Kirschner, D.A., Hollingshead, C.J., Thaxton, C., Caspar, D.L., Goodenough, D.A., 1979. Structural states of myelin observed by x-ray diffraction and freeze-fracture electron microscopy. J. Cell Biol. 82, 140–149. https://doi.org/10.1083/jcb.82.1.140 Klbik, I., Čechová, K., Milovská, S., Rusnák, J., Vlasáč, J., Melicherčík, M., Mat’ko, I., Lakota, J., Šauša, O., 2022. Cryoprotective Mechanism of DMSO Induced by the Inhibitory Effect on Eutectic NaCl Crystallization. J. Phys. Chem. Lett. 13, 11153–11159. https://doi.org/10.1021/acs.jpclett.2c03003 Korogod, N., Petersen, C.C., Knott, G.W., 2015. Ultrastructural analysis of adult mouse neocortex comparing aldehyde perfusion with cryo fixation. eLife 4, e05793. https://doi.org/10.7554/eLife.05793 Krassner, M.M., Kauffman, J., Sowa, A., Cialowicz, K., Walsh, S., Farrell, K., Crary, J.F., McKenzie, A.T., 2023. Postmortem changes in brain cell structure: a review. Free Neuropathol. 4, 4–10. https://doi.org/10.17879/freeneuropathology-2023-4790 Kumarasami, R., Verma, R., Pandurangan, K., Ramesh, J.J., Pandidurai, S., Savoia, S., Jayakumar, J., Bota, M., Mitra, P., Joseph, J., Sivaprakasam, M., 2023. A technology platform for standardized cryoprotection and freezing of large-volume brain tissues for high-resolution histology. Front. Neuroanat. 17, 1292655. https://doi.org/10.3389/fnana.2023.1292655 Landis, D.M., Reese, T.S., 1983. Cytoplasmic organization in cerebellar dendritic spines. J. Cell Biol. 97, 1169–1178. https://doi.org/10.1083/jcb.97.4.1169 Landis, D.M., Reese, T.S., 1981. Astrocyte membrane structure: changes after circulatory arrest. J. Cell Biol. 88, 660–663. https://doi.org/10.1083/jcb.88.3.660 Lee, E., Baiz, C.R., 2022. How cryoprotectants work: hydrogen-bonding in low-temperature vitrified solutions. Chem. Sci. 13, 9980–9984. https://doi.org/10.1039/D2SC03188D Lee, R.E., Allenspach, A.L., Collins, S.D., 1997. Ultrastructural effects of lethal freezing on brain, muscle and Malpighian tubules from freeze-tolerant larvae of the gall fly, Eurosta solidaginis. J. Insect Physiol. 43, 39–45. Lemler, J., Harris, S.B., Platt, C., Huffman, T.M., 2004. The arrest of biological time as a bridge to engineered negligible senescence. Ann. N. Y. Acad. Sci. 1019, 559–563. https://doi.org/10.1196/annals.1297.104 Leonard, A., Vink, R., Byard, R.W., 2016. Brain Fluid Content Related to Body Position and Postmortem Interval - An Animal Model. J. Forensic Sci. 61, 671–673. https://doi.org/10.1111/1556-4029.13038 Li, J.Z., Meng, F., Tsavaler, L., Evans, S.J., Choudary, P.V., Tomita, H., Vawter, M.P., Walsh, D., Shokoohi, V., Chung, T., Bunney, W.E., Jones, E.G., Akil, H., Watson, S.J., Myers, R.M., 2007. Sample matching by inferred agonal stress in gene expression analyses of the brain. BMC Genomics 8, 336. https://doi.org/10.1186/1471-2164-8-336 Li, J.Z., Vawter, M.P., Walsh, D.M., Tomita, H., Evans, S.J., Choudary, P.V., Lopez, J.F., Avelar, A., Shokoohi, V., Chung, T., Mesarwi, O., Jones, E.G., Watson, S.J., Akil, H., Bunney, W.E., Jr, Myers, R.M., 2004. Systematic changes in gene expression in postmortem human brains associated with tissue pH and terminal medical conditions. Hum. Mol. Genet. 13, 609–616. https://doi.org/10.1093/hmg/ddh065 Li, W., Chen, Q., Wang, X., Chen, Z., 2023. Effect of Freezing on Soybean Protein Solution. Foods Basel Switz. 12, 2650. https://doi.org/10.3390/foods12142650 Linner, J.G., Livesey, S.A., Harrison, D.S., Steiner, A.L., 1986. A new technique for removal of amorphous phase tissue water without ice crystal damage: a preparative method for ultrastructural analysis and immunoelectron microscopy. J. Histochem. Cytochem. Off. J. Histochem. Soc. 34, 1123–1135. https://doi.org/10.1177/34.9.2426340 Marshall, W.H., 1940. An Application of the Frozen Sectioning Technic for Cutting Serial Sections Thru the Brain. Stain Technol. 15, 133–138. https://doi.org/10.3109/10520294009110338 Massie, I., Selden, C., Hodgson, H., Fuller, B., 2013. Storage temperatures for cold-chain delivery in cell therapy: a study of alginate-encapsulated liver cell spheroids stored at -80 °c or -170 °c for up to 1 year. Tissue Eng. Part C Methods 19, 189–195. https://doi.org/10.1089/ten.TEC.2012.0307 Mazur, P., Leibo, S.P., Chu, E.H., 1972. A two-factor hypothesis of freezing injury. Evidence from Chinese hamster tissue-culture cells. Exp. Cell Res. 71, 345–355. https://doi.org/10.1016/0014-4827(72)90303-5 McFadden, W.C., Walsh, H., Richter, F., Soudant, C., Bryce, C.H., Hof, P.R., Fowkes, M., Crary, J.F., McKenzie, A.T., 2019. Perfusion fixation in brain banking: a systematic review. Acta Neuropathol. Commun. 7, 146. https://doi.org/10.1186/s40478-019-0799-y McGrath, J.J., 1985. Preservation of Biological Material by Freezing and Thawing, in: Shitzer, A., Eberhart, R.C. (Eds.), Heat Transfer in Medicine and Biology: Analysis and Applications. Volume 2. Springer US, Boston, MA, pp. 185–238. https://doi.org/10.1007/978-1-4684-8285-0_5 McIntyre, J.A., Gilula, N.B., Karnovsky, M.J., 1974. Cryoprotectant-induced redistribution of intramembranous particles in mouse lymphocytes. J. Cell Biol. 60, 192–203. https://doi.org/10.1083/jcb.60.1.192 McIntyre, R.L., Fahy, G.M., 2015. Aldehyde-stabilized cryopreservation. Cryobiology 71, 448–458. https://doi.org/10.1016/j.cryobiol.2015.09.003 McKee, A.C., 1999. Brain banking: basic science methods. Alzheimer Dis. Assoc. Disord. 13 Suppl 1, S39-44. McKenzie, A.T., Woodoff-Leith, E., Dangoor, D., Cervera, A., Ressler, H., Whitney, K., Dams-O’Connor, K., Wu, Z., Hillman, E.M.C., Seifert, A.C., Crary, J.F., 2022. Ex situ perfusion fixation for brain banking: a technical report. Free Neuropathol. 3, 22–22. https://doi.org/10.17879/freeneuropathology-2022-4368 Meissner, D.H., Schwarz, H., 1990. Improved cryoprotection and freeze-substitution of embryonic quail retina: a TEM study on ultrastructural preservation. J. Electron Microsc. Tech. 14, 348–356. https://doi.org/10.1002/jemt.1060140410 Meneghel, J., Kilbride, P., Morris, J.G., Fonseca, F., 2019. Physical events occurring during the cryopreservation of immortalized human T cells. PloS One 14, e0217304. https://doi.org/10.1371/journal.pone.0217304 Menz, L.J., 1971. Structural changes and impairment of function associated with freezing and thawing in muscle, nerve, and leucocytes. Cryobiology 8, 1–13. https://doi.org/10.1016/0011-2240(71)90102-7 Meyronet, D., Dorey, A., Massoma, P., Rey, C., Alix, E., Silva, K., Perrin, C., Quadrio, I., Perret-Liaudet, A., Streichenberger, N., Thomasset, N., Honnorat, J., Arzberger, T., Kretzschmar, H., 2015. The workflow from post-mortem human brain sampling to cell microdissection: a Brain Net Europe study. J. Neural Transm. Vienna Austria 1996 122, 975–991. https://doi.org/10.1007/s00702-015-1378-4 Möbius, W., Cooper, B., Kaufmann, W.A., Imig, C., Ruhwedel, T., Snaidero, N., Saab, A.S., Varoqueaux, F., 2010. Electron microscopy of the mouse central nervous system. Methods Cell Biol. 96, 475–512. https://doi.org/10.1016/S0091-679X(10)96020-2 Moor, H., Bellin, G., Sandri, C., Akert, K., 1980. The influence of high pressure freezing on mammalian nerve tissue. Cell Tissue Res. 209, 201–216. https://doi.org/10.1007/BF00237626 Murray, K.A., Gibson, M.I., 2022. Chemical approaches to cryopreservation. Nat. Rev. Chem. 6, 579–593. https://doi.org/10.1038/s41570-022-00407-4 Nochlin, D., Mackenzie, A.P., Bryant, E.M., Norwood, T.H., Sumi, S.M., 1993. A simple method of rapid freezing adequately preserves brain tissue for immunocytochemistry, light and electron microscopic examination. Acta Neuropathol. (Berl.) 86, 645–650. https://doi.org/10.1007/BF00294305 Notter, T., Panzanelli, P., Pfister, S., Mircsof, D., Fritschy, J.-M., 2014. A protocol for concurrent high-quality immunohistochemical and biochemical analyses in adult mouse central nervous system. Eur. J. Neurosci. 39, 165–175. https://doi.org/10.1111/ejn.12447 Okotrub, K.A., Surovtsev, N.V., 2013. Raman scattering evidence of hydrohalite formation on frozen yeast cells. Cryobiology 66, 47–51. https://doi.org/10.1016/j.cryobiol.2012.11.002 Otubo, A., Maejima, S., Oti, T., Satoh, K., Ueda, Y., Morris, J.F., Sakamoto, T., Sakamoto, H., 2021. Immunoelectron Microscopic Characterization of Vasopressin-Producing Neurons in the Hypothalamo-Pituitary Axis of Non-Human Primates by Use of Formaldehyde-Fixed Tissues Stored at -25 °C for Several Years. Int. J. Mol. Sci. 22, 9180. https://doi.org/10.3390/ijms22179180 Ozyoruk, K.B., Can, S., Darbaz, B., Başak, K., Demir, D., Gokceler, G.I., Serin, G., Hacisalihoglu, U.P., Kurtuluş, E., Lu, M.Y., Chen, T.Y., Williamson, D.F.K., Yılmaz, F., Mahmood, F., Turan, M., 2022. A deep-learning model for transforming the style of tissue images from cryosectioned to formalin-fixed and paraffin-embedded. Nat. Biomed. Eng. 6, 1407–1419. https://doi.org/10.1038/s41551-022-00952-9 Paljärvi, L., Garcia, J.H., Kalimo, H., 1979. The efficiency of aldehyde fixation for electron microscopy: stabilization of rat brain tissue to withstand osmotic stress. Histochem. J. 11, 267–276. https://doi.org/10.1007/BF01005026 Paynter, S.J., 2008. Principles and practical issues for cryopreservation of nerve cells. Brain Res. Bull. 75, 1–14. https://doi.org/10.1016/j.brainresbull.2007.08.004 Pegg, D.E., 2007. Principles of cryopreservation. Methods Mol. Biol. Clifton NJ 368, 39–57. https://doi.org/10.1007/978-1-59745-362-2_3 Pegg, D.E., 1976. Long-term preservation of cells and tissues: a review. J. Clin. Pathol. 29, 271–285. Pérez-Bermúdez, I., Castillo-Suero, A., Cortés-Inostroza, A., Jeldrez, C., Dantas, A., Hernández, E., Orellana-Palma, P., Petzold, G., 2023. Observation and Measurement of Ice Morphology in Foods: A Review. Foods Basel Switz. 12, 3987. https://doi.org/10.3390/foods12213987 Peters, S., 2010. A Practical Guide to Frozen Section Technique, A Practical Guide to Frozen Section Technique. https://doi.org/10.1007/978-1-4419-1234-3 Petzold, G., Aguilera, J.M., 2009. Ice Morphology: Fundamentals and Technological Applications in Foods. Food Biophys. 4, 378–396. https://doi.org/10.1007/s11483-009-9136-5 Pichugin, Y., Fahy, G.M., Morin, R., 2006. Cryopreservation of rat hippocampal slices by vitrification. Cryobiology 52, 228–240. https://doi.org/10.1016/j.cryobiol.2005.11.006 Pontén, U., Ratcheson, R.A., Siesjö, B.K., 1973. Metabolic changes in the brains of mice frozen in liquid nitrogen. J. Neurochem. 21, 1211–1216. Priemer, D.S., Wysozan, T., Zahedi, F., Alrabadi, N., Mesa, H., Vortmeyer, A.O., 2023. Qualitative Comparison of Cryostat- versus Snap-Frozen Neurosurgical Intraoperative Consultations. Int. J. Surg. Pathol. 31, 949–956. https://doi.org/10.1177/10668969221117987 Rabin, Y., Steif, P.S., Hess, K.C., Jimenez-Rios, J.L., Palastro, M.C., 2006. Fracture formation in vitrified thin films of cryoprotectants. Cryobiology 53, 75–95. https://doi.org/10.1016/j.cryobiol.2006.03.013 Ramirez, E.P.C., Keller, C.E., Vonsattel, J.P., 2018. The New York Brain Bank of Columbia University: practical highlights of 35 years of experience. Handb. Clin. Neurol. 150, 105–118. https://doi.org/10.1016/B978-0-444-63639-3.00008-6 Reger, J.F., Escaig, F., 1988. A comparative fine structure study of rat cerebral cortex following ultra-rapid freezing and conventional chemical fixation procedures. J. Submicrosc. Cytol. Pathol. 20, 691–700. Robards, A.W., Sleytr, U.B., 1985. Low Temperature Methods in Biological Electron Microscopy. Elsevier Science. Rosene, D.L., Rhodes, K.J., 1990. Cryoprotection and Freezing Methods to Control Ice Crystal Artifact in Frozen Sections of Fixed and Unfixed Brain Tissue, in: Conn, P.M. (Ed.), Methods in Neurosciences, Quantitative and Qualitative Microscopy. Academic Press, pp. 360–385. https://doi.org/10.1016/B978-0-12-185255-9.50026-2 Rosene, D.L., Roy, N.J., Davis, B.J., 1986. A cryoprotection method that facilitates cutting frozen sections of whole monkey brains for histological and histochemical processing without freezing artifact. J. Histochem. Cytochem. Off. J. Histochem. Soc. 34, 1301–1315. https://doi.org/10.1177/34.10.3745909 Rossner, M.J., Hirrlinger, J., Wichert, S.P., Boehm, C., Newrzella, D., Hiemisch, H., Eisenhardt, G., Stuenkel, C., von Ahsen, O., Nave, K.-A., 2006. Global transcriptome analysis of genetically identified neurons in the adult cortex. J. Neurosci. Off. J. Soc. Neurosci. 26, 9956–9966. https://doi.org/10.1523/JNEUROSCI.0468-06.2006 Rubinsky, B., Pegg, D.E., 1988. A mathematical model for the freezing process in biological tissue. Proc. R. Soc. Lond. B Biol. Sci. 234, 343–358. https://doi.org/10.1098/rspb.1988.0053 Ryan, K.P., 1991. Rapid cryogenic fixation of biological specimens for electron microscopy. Schäfer, A.T., Kaufmann, J.D., 1999. What happens in freezing bodies? Experimental study of histological tissue change caused by freezing injuries. Forensic Sci. Int. 102, 149–158. https://doi.org/10.1016/s0379-0738(99)00043-2 Seilhean, D., Gansmüller, A., Baron-Van Evercooren, A., Gumpel, M., Lachapelle, F., 1996. Myelination by transplanted human and mouse central nervous system tissue after long-term cryopreservation. Acta Neuropathol. (Berl.) 91, 82–88. https://doi.org/10.1007/s004010050396 Senda, T., Iizuka-Kogo, A., Shimomura, A., 2005. Visualization of the nuclear lamina in mouse anterior pituitary cells and immunocytochemical detection of lamin A/C by quick-freeze freeze-substitution electron microscopy. J. Histochem. Cytochem. Off. J. Histochem. Soc. 53, 497–507. https://doi.org/10.1369/jhc.4A6478.2005 Shabihkhani, M., Lucey, G.M., Wei, B., Mareninov, S., Lou, J.J., Vinters, H.V., Singer, E.J., Cloughesy, T.F., Yong, W.H., 2014. The procurement, storage, and quality assurance of frozen blood and tissue biospecimens in pathology, biorepository, and biobank settings. Clin. Biochem. 47, 258–266. https://doi.org/10.1016/j.clinbiochem.2014.01.002 Shepherd, C.E., Alvendia, H., Halliday, G.M., 2019. Brain Banking for Research into Neurodegenerative Disorders and Ageing. Neurosci. Bull. 35, 283–288. https://doi.org/10.1007/s12264-018-0326-3 Siksou, L., Rostaing, P., Lechaire, J.-P., Boudier, T., Ohtsuka, T., Fejtová, A., Kao, H.-T., Greengard, P., Gundelfinger, E.D., Triller, A., Marty, S., 2007. Three-dimensional architecture of presynaptic terminal cytomatrix. J. Neurosci. Off. J. Soc. Neurosci. 27, 6868–6877. https://doi.org/10.1523/JNEUROSCI.1773-07.2007 Sosinsky, G.E., Crum, J., Jones, Y.Z., Lanman, J., Smarr, B., Terada, M., Martone, M.E., Deerinck, T.J., Johnson, J.E., Ellisman, M.H., 2008. The combination of chemical fixation procedures with high pressure freezing and freeze substitution preserves highly labile tissue ultrastructure for electron tomography applications. J. Struct. Biol. 161, 359–371. https://doi.org/10.1016/j.jsb.2007.09.002 Studer, D., Zhao, S., Chai, X., Jonas, P., Graber, W., Nestel, S., Frotscher, M., 2014. Capture of activity-induced ultrastructural changes at synapses by high-pressure freezing of brain tissue. Nat. Protoc. 9, 1480–1495. https://doi.org/10.1038/nprot.2014.099 Suda, I., Kito, K., Adachi, C., 1966. Viability of long term frozen cat brain in vitro. Nature 212, 268–270. https://doi.org/10.1038/212268a0 Szabo, G.V., Burg, T.P., 2024. Time Resolved Cryo-Correlative Light and Electron Microscopy. Adv. Funct. Mater. 34, 2313705. https://doi.org/10.1002/adfm.202313705 Taqi, S.A., Sami, S.A., Sami, L.B., Zaki, S.A., 2018. A review of artifacts in histopathology. J. Oral Maxillofac. Pathol. JOMFP 22, 279. https://doi.org/10.4103/jomfp.JOMFP_125_15 Tian, M., Reger, J.F., Armstrong, W.E., 1991. Electron microscopic and immunocytochemical study of rapidly frozen, freeze-substituted neural lobes of rats. J. Neurocytol. 20, 79–96. https://doi.org/10.1007/BF01279613 Tofte, K., Berger, C., Torp, S.H., Solheim, O., 2014. The diagnostic properties of frozen sections in suspected intracranial tumors: A study of 578 consecutive cases. Surg. Neurol. Int. 5, 170. https://doi.org/10.4103/2152-7806.146153 Tourtellotte, W.W., Rosario, I.P., Conrad, A., Syndulko, K., 1993. Human neuro-specimen banking 1961-1992. The National Neurological Research Specimen Bank (a donor program of pre- and post-mortem tissues and cerebrospinal fluid/blood; and a collection of cryopreserved human neurological specimens for neuroscientists). J. Neural Transm. Suppl. 39, 5–15. Trump, B.F., Young, D.E., Arnold, E.A., Stowell, R.E., 1965. EFFECTS OF FREEZING AND THAWING ON THE STRUCTURE, CHEMICAL CONSTITUTION, AND FUNCTION OF CYTOPLASMIC STRUCTURES. Fed. Proc. 24, S144-168. Ulfig, N., Nickel, J., Bohl, J., 1998. Monoclonal antibodies SMI 311 and SMI 312 as tools to investigate the maturation of nerve cells and axonal patterns in human fetal brain. Cell Tissue Res. 291, 433–443. https://doi.org/10.1007/s004410051013 van der Wijngaart, H., Jagga, S., Dekker, H., de Goeij, R., Piersma, S.R., Pham, T.V., Knol, J.C., Zonderhuis, B.M., Holland, H.J., Jiménez, C.R., Verheul, H.M.W., Vanapalli, S., Labots, M., 2023. Advancing wide implementation of precision oncology: A liquid nitrogen-free snap freezer preserves molecular profiles of biological samples. Cancer Med. 12, 10979–10989. https://doi.org/10.1002/cam4.5781 Vanhecke, D., Graber, W., Studer, D., 2010. Rapidly excised and cryofixed rat tissue. Methods Cell Biol. 96, 513–527. https://doi.org/10.1016/S0091-679X(10)96021-4 Vonsattel, J.P., Aizawa, H., Ge, P., DiFiglia, M., McKee, A.C., MacDonald, M., Gusella, J.F., Landwehrmeyer, G.B., Bird, E.D., Richardson, E.P., 1995. An improved approach to prepare human brains for research. J. Neuropathol. Exp. Neurol. 54, 42–56. https://doi.org/10.1097/00005072-199501000-00006 Vonsattel, J.P.G., Amaya, M. del P., Cortes, E.P., Mancevska, K., Keller, C.E., 2008. 21st century brain banking practical prerequisites and lessons from the past: the experience of New York Brain Bank – Taub Institute - Columbia University. Cell Tissue Bank. 9, 247–258. https://doi.org/10.1007/s10561-008-9079-y Watson, R.E.J., Wiegand, S.J., Clough, R.W., Hoffman, G.E., 1986. Use of cryoprotectant to maintain long-term peptide immunoreactivity and tissue morphology. Peptides 7, 155–159. https://doi.org/10.1016/0196-9781(86)90076-8 Weng, L., Stott, S.L., Toner, M., 2019. Exploring Dynamics and Structure of Biomolecules, Cryoprotectants, and Water Using Molecular Dynamics Simulations: Implications for Biostabilization and Biopreservation. Annu. Rev. Biomed. Eng. 21, 1–31. https://doi.org/10.1146/annurev-bioeng-060418-052130 Wieser, M., Burger, S., Ertl, R., Kummer, S., Stargardt, M., Walter, I., 2022. Example for process validation in biobanking: Fit for purpose testing of a cryopreservation method without isopentane. Front. Mol. Biosci. 9, 876670. https://doi.org/10.3389/fmolb.2022.876670 Wong, G., Greenhalgh, T., Westhorp, G., Buckingham, J., Pawson, R., 2013. RAMESES publication standards: realist syntheses. BMC Med. 11, 21. https://doi.org/10.1186/1741-7015-11-21 Woods, E.J., Mullen, S.F., 2016. Organ Preservation: Cryobiology and Beyond. Curr. Stem Cell Rep. 2, 104–117. https://doi.org/10.1007/s40778-016-0042-8 Wowk, B., 2010. Thermodynamic aspects of vitrification. Cryobiology 60, 11–22. https://doi.org/10.1016/j.cryobiol.2009.05.007 Wowk, B., Phan, J., Pagotan, R., Galvez, E., Fahy, G.M., 2024. 27 MHz constant field dielectric warming of kidneys cryopreserved by vitrification. Cryobiology 104893. https://doi.org/10.1016/j.cryobiol.2024.104893 Zennoune, A., Latil, P., Flin, F., Perrin, J., Weitkamp, T., Scheel, M., Geindreau, C., Benkhelifa, H., Ndoye, F.-T., 2022. Investigating the influence of freezing rate and frozen storage conditions on a model sponge cake using synchrotron X-rays micro-computed tomography. Food Res. Int. Ott. Ont 162, 112116. https://doi.org/10.1016/j.foodres.2022.112116 Zhang, M., Oldenhof, H., Sieme, H., Wolkers, W.F., 2016. Freezing-induced uptake of trehalose into mammalian cells facilitates cryopreservation. Biochim. Biophys. Acta 1858, 1400–1409. https://doi.org/10.1016/j.bbamem.2016.03.020 Zhao, S., Studer, D., Graber, W., Nestel, S., Frotscher, M., 2012. Fine structure of hippocampal mossy fiber synapses following rapid high-pressure freezing. Epilepsia 53 Suppl 1, 4–8. https://doi.org/10.1111/j.1528-1167.2012.03469.x Zilles, K., Schleicher, A., Palomero-Gallagher, N., Amunts, K., 2002. 21 - Quantitative Analysis of Cyto- and Receptor Architecture of the Human Brain, in: Toga, A.W., Mazziotta, J.C. (Eds.), Brain Mapping: The Methods (Second Edition). Academic Press, San Diego, pp. 573–602. https://doi.org/10.1016/B978-012693019-1/50023-X Zuber, B., Nikonenko, I., Klauser, P., Muller, D., Dubochet, J., 2005. The mammalian central nervous synaptic cleft contains a high density of periodically organized complexes. Proc. Natl. Acad. Sci. 102, 19192–19197. https://doi.org/10.1073/pnas.0509527102
Copyright: © 2024 The author(s). This is an open access article distributed under the terms of the Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited, a link to the Creative Commons license is provided, and any changes are indicated. The Creative Commons Public Domain Dedication waiver (https://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated. |