|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Free Neuropathology 5:14 (2024) |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Original Paper |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Stem cell-associated transcription factors in non-functioning pituitary neuroendocrine tumours |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Kristin Astrid Øystese1, Nicoleta Cristina Olarescu1,2, Cecilia Lindskog3, Fabjola Xheka4,5, Jon Berg-Johnsen2,6, Jens Petter Berg2,7, Jens Bollerslev1,2, Olivera Casar-Borota4,8 |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Corresponding author: |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Additional resources and electronic supplementary material: supplementary material |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Submitted: 29 February 2024 |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Keywords: Pituitary neoplasm, Non-functioning pituitary adenoma, Neoplastic stem cells, Stem cells |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Abstract
Background: Cells with stem cell features have been described in pituitary neuroendocrine tumours (PitNETs). Transcription factors SOX2 and SOX9 are stem cell-associated markers while the pituitary progenitor marker PROP1 is involved in anterior pituitary development. We characterised the presence of these markers known to be present in the human pituitary in non-functioning (NF) PitNETs. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Introduction The pituitary gland is the main regulator of the endocrine systems, and has a complex development and maturation during life. The pituitary orchestrates growth, metabolism and reproduction in humans, and adapts to major physiological life changes such as puberty, pregnancy and menopause. The different cell lineages of the anterior pituitary lobe are derived from Rathke's pouch during embryogenesis [1]. Rathke's pouch contains stem cells and progenitor cells expressing transcription factors, e.g. SRY-Box Transcription Factor 2 (SOX2), SRY-Box Transcription Factor 9 (SOX9) and Prophet of PIT1 (PROP1) [2]. Single cell sequencing shows that SOX2, SOX9 and PROP1 are expressed in stem cell populations in the human fetal pituitary [3]. Adult stem cells have been identified in several tissues including the pituitary, and have the capability of self-renewal, and the ability to differentiate in specific cell types that are typically restricted to the tissue of origin [3, 4]. The pituitary adaptation to different life events might be facilitated by the presence of stem cells [5]. PROP1 is a transcription factor found in the early development of the pituitary gland and is known to be expressed in pituitary stem cells [3]. PROP1 is necessary for the development of the PIT1 and SF1 lineages. Moreover, PROP1 has a role in the migration of developing pituitary cells from Rathke's pouch in early pituitary development [6–8]. For simplicity, PROP1 will be addressed as a stem cell-associated marker throughout the manuscript together with SOX2 and SOX9. Pituitary neuroendocrine tumours (PitNETs) are common intracranial neoplasms of epithelial origin. They are classified immunohistochemically based on the staining for the different adenohypophysial hormones FSH, LH, ACTH, GH, Prolactin and TSH and the pituitary transcription factors SF1, TPIT and PIT1 [9]. Further, the tumours are clinically characterised by their functionality. Approximately half of the PitNETs are clinically non-functioning, lacking clinical features and symptoms related to hormone overproduction [10, 11]. The majority of non-functioning PitNETs (NF-PitNETs) are gonadotroph (SF1) in origin, followed by corticotroph (TPIT) and rarely by PIT1 cell lineage tumours [12, 13]. The NF-PitNETs are usually detected due to compression symptoms from the surrounding structures, mainly yielding symptoms from the visual apparatus or from hypopituitarism. However, an increasing number of tumours are detected incidentally by imaging procedures taken for unrelated reasons [14]. To date there is no medical treatment available for the NF-PitNETs. Tumour-directed treatment involves pituitary surgery. A substantial proportion of patients presents with postoperative tumour remnants that may grow, and there are no reliable markers differentiating the tumours needing adjuvant or repeated intervention. The tumourigenesis of PitNETs is poorly understood. Most tumours are sporadic, and only a small proportion is associated with gene mutations e.g. in AIP or MEN [15]. In a rat experimental pituitary tumour model, the level of SOX9 was decreased at the initial stage of tumourigenesis; however both SOX2 and SOX9 expression levels were similar to those observed in normal pituitary gland at a more advanced stage of tumourigenesis [16]. Cells with stem cell-like features have been found in histological specimens from human adenohypophysial tumours [17] and single cell mRNA-sequencing has shown clustering of pituitary stem cell markers in both normal and tumourous pituitary stem cells [18]. SOX2 has been found in approximately 20 % of human NF-PitNETs [19]. Furthermore, nuclear coexpression of PROP1 and SOX2 was found in tumour stem cells isolated from NF-PitNETs [20]. It has been hypothesised that pituitary stem cells play a role in the development of PitNETs by paracrine effects, while tumour models have shown that the stem cells themselves do not give rise to tumour cells [21, 22]. Yet there is still a substantial knowledge gap concerning the role of tumour stem cells at different stages of tumourigenesis. In this study, we characterised the presence of stem cell-associated transcription factors known to be present in the adult human pituitary in NF-PitNETs and correlated them to clinical and molecular markers. Methods Patients The study retrospectively included patients operated for clinical NF-PitNET between 1998 and 2009, where tumour tissue from primary pituitary surgery was available for immunohistochemical (IHC) subclassification (N = 158). All tumours were clinically classified as non-functioning at the time of surgery. Seventeen patients lacked tumour tissue available for IHC characterisation or frozen tumour tissue available for RT-qPCR (9 SF1, 4 null-cell, 3 TPIT and 1 PIT1 tumour), leaving 141 patients (129 tumour samples available for IHC analyses and 81 for qRT-PCR) for analysis of the stem cell-associated markers. Additionally four null-cell NF-PitNETs (not staining for anterior pituitary hormones or transcription factors) were excluded from the analyses due to lack of consistent IHC characterisation. All patients were operated at the same tertiary referral centre. Data from magnetic resonance imaging (MRI) were available for 60 patients (46 with gonadotroph, 9 with corticotroph and 5 with PIT1 tumours). Tumour volume was measured by the Cavalieri method, as previously described [23], and invasion was defined by the Knosp criteria. A tumour with a Knosp grade ≥ 3 on either side was defined as an invasive tumour [24, 25]. Repeated surgery or radiation therapy less than 12 months after primary surgery was defined as adjuvant treatment to the primary surgery. Hence, reintervention was defined as surgery or radiation therapy more than 12 months after primary surgery. Informed consent was obtained from all living patients. The regional ethics committee (REK 2014/635) and the hospital authority approved the study. Histopathology and Immunohistochemistry The original hematoxylin and eosin stained sections from all tumours were reviewed by a pathologist (OCB) to confirm the presence of representative tumour tissue. Tissue microarrays (TMAs) were constructed containing two replicate 1 mm cores from formalin-fixed paraffin-embedded (FFPE) tissue samples of representative areas [26]. TMA blocks were sectioned, mounted on adhesive slides and dried at room temperature overnight, followed by baking at 50 °C for 12–24 h. The sections were deparaffinised in xylene, hydrated in graded alcohols and blocked for endogenous peroxidase in 0.3 % hydrogen peroxide diluted in 95 % ethanol. For antigen retrieval, slides were boiled for 4 min at 125 °C in citrate buffer pH6 (Dako Target Retrieval Solution (Agilent Technologies, Santa Clara, CA) using a Decloaking chamber (Biocare Medical, Walnut Creek, CA). Automated immunohistochemistry for SOX2, SOX9 and PROP1 was performed essentially as previously described [27, 28] using an Autostainer 480 instrument (Thermo Fischer Scientific, Waltham, MA). Primary antibodies against SOX2 (371R-15, Cell Marque, Rocklin, CA), SOX9 (AMAb90795, Atlas Antibodies, Bromma, Sweden) and PROP1 (HPA049839, Atlas Antibodies) were diluted 1 : 50, 1 : 150 or 1 : 1500, respectively, in UltraAb Diluent (Thermo Fisher Scientific) followed by incubation for 30 min at room temperature. The slides were further incubated with the secondary reagent anti-rabbit/mouse horseradish peroxidase-conjugated UltraVision (Thermo Fischer Scientific) for 30 min at room temperature, and developed for 10 min using Diaminobenzidine (DAB) Quanto (Thermo Fisher Scientific) as chromogen. All incubations were followed by rinse in wash buffer (Thermo Fisher Scientific) twice for 5 min. Slides were counterstained in Mayer's hematoxylin (Histolab, Gothenburg, Sweden) and coverslipped using Pertex (Histolab) as mounting medium. The slides stained for SOX2, SOX9 and PROP1 were scanned in Hamamatsu Nanozoomer S60 at 40 x magnification. The microphotographs illustrating the immunohistochemical results were taken from the scanned images. Multiplex Immunofluorescence (mIF) Slides were baked overnight at 50 °C, deparaffinised in xylene and rehydrated in graded alcohols (99.9 %, 95 %, and 80 %) down to deionised water. Endogenous peroxidase was blocked using 0.3 % hydrogen peroxide in 95 % alcohol, and heat-induced epitope retrieval (HIER) was performed in a decloaking chamber (Biocare Medical) at 125 °C for 4 min while the slides were immersed in 1X Target Retrieval Solution pH 6.0 (Agilent Technologies Inc., Santa Clara, CA, USA). Slides were then cooled to approximately 90 °C before rinsing with deionised water. After HIER the slides were treated with a LED-light bleaching process immersed in a bleaching solution consisting of 0.2M glycine, 1.5 % hydrogen peroxide, and 1X TBS+Tween (TA-999-TT, Thermo Fisher Scientific) for 1 h in RT. Slides were incubated using a multiplex panel of six antibodies in a 6-cycle antibody staining process with intermediary elution steps after each cycle. Antibody stripping/elution was performed by boiling the slides in a decloaking chamber (Biocare Medical) at 90 °C for 20 min and slides immersed in 1X Target Retrieval Solution, pH 6.0 (Agilent Technologies). Full staining cycle information (panel antibody markers, dilutions, reagents, incubation times, OPAL fluorophores) are available in supplementary table 1. One cycle of staining included blocking, primary antibody incubation, anti-rabbit IgG (H+L) with horseradish peroxidase (HRP) polymer, and an OPAL fluorophore (Akoya Biosciences, Marlborough, MA, USA). All cycles were performed at room temperature (RT) using the Austostainer 480S (Thermo Fischer Scientific) and HRP-kit from Epredia (Epredia Ultravision LP HRP-kit, Breda, Netherlands). After the last cycle, slides were incubated with the OPAL 780 fluorophore-conjugated anti-DIG antibody and 4',6-diamidino-2-phenylindole (DAPI) (Invitrogen™, D1306, Thermo Fisher Scientific). Slides were then mounted using Prolong™ Glass Antifade mounting media and left overnight at RT after which they were digitalised at 40 x magnification using PhenoImager (Akoya Biosciences). Spectral unmixing and export of images were performed using the built-in spectral library of the inForm software (Akoya Biosciences). Multiplex IHC was performed on one TMA including cores from 50 NF-PitNETs of different types. Classification of immunohistochemically stained samples The immunohistochemical classification was based on the expression of the anterior pituitary lobe hormones FSHβ, LHβ, ACTH, GH, PRL, TSH and alpha subunit and the transcription factors SF1, TPIT and PIT1 as previously described [13, 29]. The tumours were classified into three groups based on their cell line of origin: gonadotroph NF-PitNETs (SF1), corticotroph NF-PitNETs (TPIT) and PIT1 NF-PitNETs. In the PIT1 group, 4 tumours stained for PRL, 1 for PRL and GH, 2 stained for alpha subunit alone, and 1 tumour was plurihormonal expressing TSH in a proportion of cells and FSHβ and/or LHβ as well as alpha subunit in scattered cells. The staining for FSHβ and LHβ was graded from 0–4 based on the percentage of positive cells (0 = no positive cells, 1 = 0–10 % positive cells, 2 = 10–50 % positive cells, 3 = 50–80 % positive cells and 4 = > 80 % positive cells). The staining for estrogen receptor α (ERα) was based on the immunoreactive score (IRS), being the product of the percentage of positive staining cells (0 = 0 %; 1 = 1–10 %; 2 = 10–50 %; 3 = 50–80 %; and 4 = > 80 %) and the predominant staining intensity (0: No staining; 1: Weak staining; 2: Moderate staining; 3: Strong staining). The stem cell-associated markers SOX2, SOX9 and PROP1 were scored based on the proportion of positively stained cells (staining score 0 = no positive cells, 1 = staining in scattered cells, 2 = staining in 1–5 % of cells, 3 = staining in 5–10 % of cells, 4 = staining in 10–30 % of cells, 5 = staining in 30–50 % of cells and 6 = staining in > 50 % of cells). All positive cells showed a distinct nuclear immunolabeling. RT-qPCR Reverse transcription (RT) was performed using a high-capacity cDNA reverse transcription kit (Applied Biosystems, Foster City, CA, USA) by a Labnet MultiGene Gradient Thermal Cycler (Labnet International Inc., Edison, NJ, USA) according to the manufacturer's protocol. After the reaction, the cDNA was diluted to a ratio of 1 : 10. Quantitative real time polymerase chain reaction (RT-qPCR) was performed in an ABI 7900 apparatus (Applied Biosystems, Foster City, CA, USA) using Power SYBR Green Master Mix (Applied Biosystems, Foster City, CA, USA). Samples were dispensed in the corresponding wells by an automated pipetting system (epMotion® 5070 CB, Hamburg, Germany). RT-qPCR was performed using a previously described protocol [30]. The primer sequences are available upon request. All RT-qPCR experiments were in accordance with the Minimum Information for Publication of Quantitative Real-Time PCR Experiments (MIQE) guidelines [31]. GAPDH or geometric mean of GAPDH and ALAS were used as housekeeping genes [30, 32]. Frozen tumour tissue was available for RT-qPCR analyses of SOX2, SOX9 and PROP1 in 66, 69 and 70 gonadotroph, three, three and four TPIT and three, three and four PIT1 samples, respectively. Statistics Chi square and Fisher's exact tests were used for group comparison of nominal data. Mann- Whitney U and Kruskall Wallis tests were used to compare continuous data between different groups. Spearman's correlation coefficient (rs) were analysed for correlation analyses. Cox regression and Kaplan-Meier analyses were used to analyse the association between reintervention and presence of SOX2. The Cox regression analysis was performed only for the gonadotroph tumours to prevent bias from the other groups with a small number of tumours. Group differences were considered statistically significant at the 5 % significance level. All statistical tests were two-sided. Statistical analyses were performed using SPSS Version 28, and figures 4 and 6 were made using GraphPad Prism 9.3.1. Results Clinical data and stem cell-associated markers Tumour tissues for RT-qPCR and/or for IHC from 137 patients with NF-PitNETs (Table 1) were eligible for analysis. Six patients (5 SF1 and 1 TPIT) had less than 12 months follow-up and were excluded from reintervention analysis. Six patients went through radiation therapy less than 12 months after primary surgery (4 SF1 and 2 TPIT); one of these went through additional reintervention.
The patients in the PIT1 group were younger than the patients in the SF1 and TPIT groups. The tumours in the PIT1 group were smaller than in the SF1 group; however preoperative MRI was only available for five of the PIT1 tumours. The subgroups were otherwise equal concerning age, gender and follow-up (Table 1). Stem cell associated-markers are expressed at a low level in NF-PitNETs The majority of NF-PitNETs were negative for SOX2, SOX9 and PROP1 (staining score 0) or positive only in scattered cells (staining score 1) (Fig 1). The staining score was concordant between replicate TMA punches from the same tumour. No tumours presented staining in more than 50 % of tumour cells (staining score 6). We found a moderate to strong correlation between the relative mRNA levels for SOX2, SOX9 and PROP1 and the IHC staining for the corresponding proteins (Table 2). Due to the large number of tumours negative (staining score 0) for the three markers, we also analysed the correlations when excluding these tumours. The correlations between mRNA and IHC levels remained significant for SOX2 and SOX9 despite a lower number of tumours (SOX2 n = 18, rs = 0.51, p = 0.03; SOX9 n = 51, rs = 0.58, p < 0.001). There was a borderline significance for PROP1, though the correlation coefficient remained at approximately the same level (PROP1 n = 15, rs = 0.51, p = 0.054). SOX2, SOX9 and PROP1 showed a moderate to strong correlation between themselves, both at the gene and the protein expression levels (Table 2).
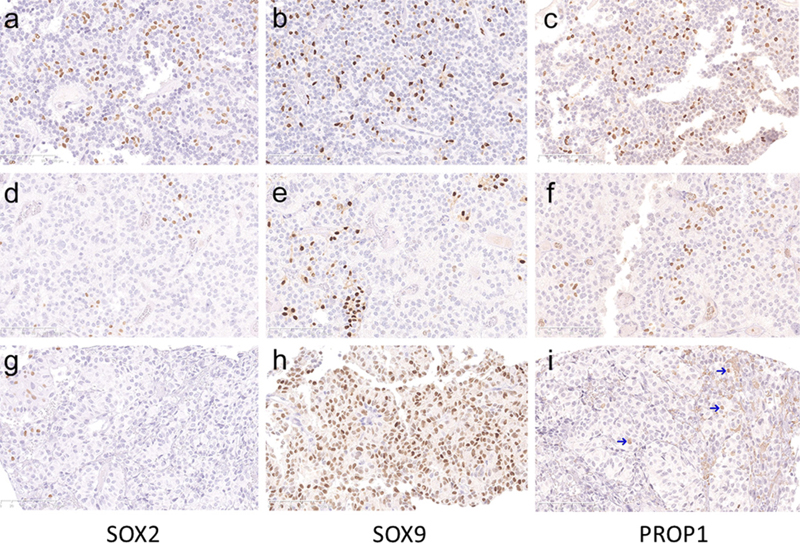
Figure 1: Two gonadotroph tumours (upper and middle row) demonstrating immunolabeling for SOX2, SOX9 and PROP1 with score 4 (a, b), score 5 (c) and score 2 (d-f). A silent corticotroph tumour (bottom row) demonstrating immunolabeling for SOX2, SOX9 and PROP1 with score 1 (g), score 5 (h) and score 1 (i). PROP1-positive cells in I are pointed with arrows to distinguish them from erythrocytes that show unspecific staining by PROP1 IHC analysis.
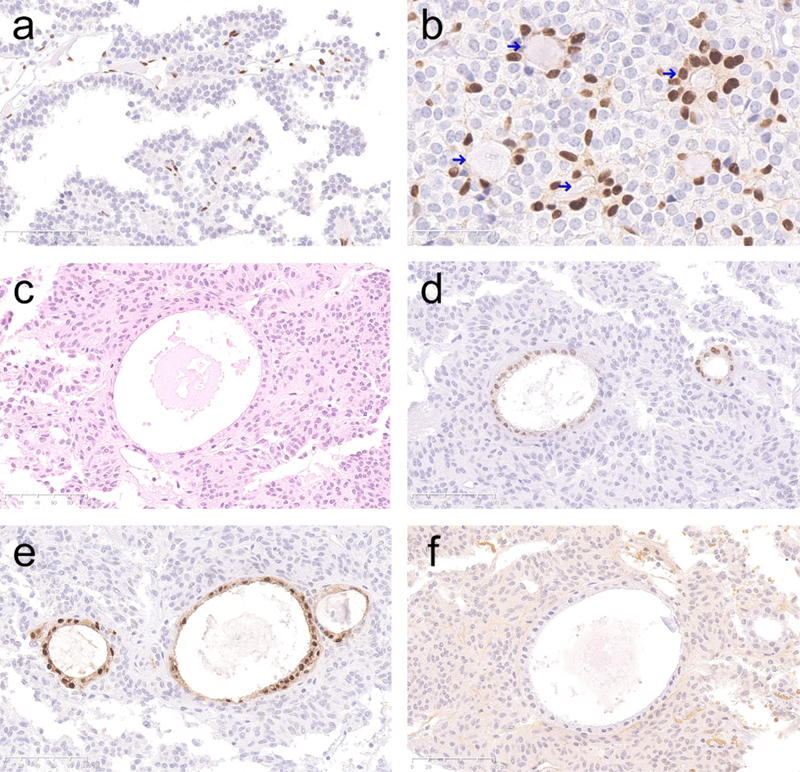
The stem cell-associated markers presented a distinct nuclear immunolabeling in positive cells. Some of these cells had a morphology similar to the tumour cells, whereas some positive cells seemed to be endothelial cells by location and morphology. Occasionally, positive cells aggregated in small groups surrounding luminal spaces. If identified within the tumour, epithelial cystic remnants demonstrated strong expression of SOX2 and SOX9 (Fig. 2). All immunolabeled cells except the epithelial cells in the cystic remnants were taken into analysis.
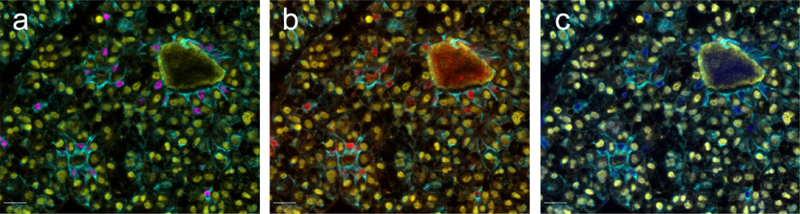
Figure 2: Immunolabeling of stem cell-associated markers in cells with a location and morphology corresponding to endothelial cells (a, SOX9, x 200) and in cells aggregating in small groups (niches, arrows) around luminal structures (b, SOX9, x 400). Epithelial cells in the remnants of Rathke's pouch within a gonadotroph tumour (c, HE) demonstrating immunolabeling for SOX2 (d) and SOX9 (e), but not PROP1 (f), (x 200). Multiplex IHC was performed on a subset of the tumours and demonstrated frequent coexpression of SOX2 and SOX9. Moreover, stem cell-associated markers were usually coexpressed with the epithelial marker. However, pituitary transcription factors, usually positive in all or the vast majority of the tumour cells, were not expressed in cells positive for stem cell-associated markers (Fig. 3).
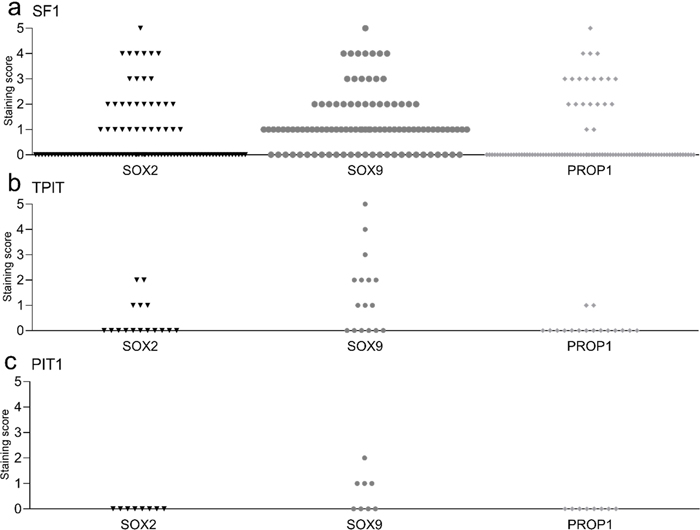
Figure 3: Multiplex IHC on a section from a gonadotroph tumour. SF1 (in yellow) and cytokeratin (in green) are illustrated on all three microphotographs. Coexpression of SOX9 (in pink) and cytokeratin is shown in a; and coexpression of SOX2 (in red) and cytokeratin is shown in b. In c (no filter for SOX2 and SOX9), a blue colour indicates nuclear staining in the cells labelled for SOX2 and SOX9 in a and b; however, no SF1 staining is seen in the nuclei of those cells. Stem cell-associated markers between different subgroups of NF-PitNETs The distribution of SOX2, SOX9 or PROP1 staining in the different subgroups of NF-PitNETs is presented in Fig. 4. There was no difference in SOX2, SOX9 or PROP1 staining between the gonadotroph (Fig. 4a) and the corticotroph (Fig. 4b) subgroups of NF-PitNETs (p = 0.91, p = 0.37 and p = 0.74, respectively). None of the tumours in the PIT1 group (Fig. 4c) (n = 8) stained for SOX2 or PROP1, while four out of eight tumours stained for SOX9, without statistical difference in comparison with the other two groups.
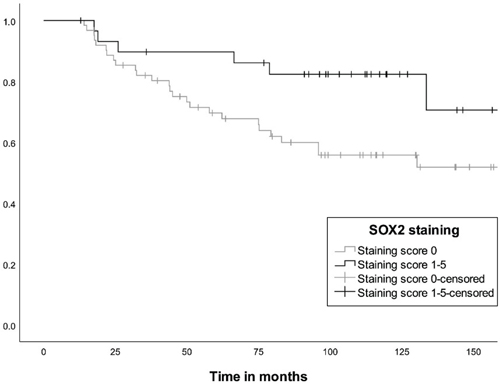
Figure 4: Distribution of stem cell-associated markers in different NF-PitNETs. Most tumours presented staining scores of 0 or 1 for SOX2 (n = 91 [64.5 %] and n = 15 [10.5 %] respectively), SOX9 (n = 41 [29 %] and n = 51 [36 %] respectively) and for PROP1 (n = 106 [75 %] and n = 4 [3 %] respectively). Proportion of positively stained cells in subgroups of NF-PitNETs. A total of 125 tumours were investigated by immunohistochemistry: 101 gonadotroph (SF1), 16 corticotroph (TPIT) and 8 from the PIT1 lineage. We did not find a significant difference in staining between the groups. Stem cell-associated markers and reintervention The tumours with no staining for SOX2 presented a higher rate of reintervention than the tumours with a SOX2 staining score ≥ 1 (n = 34 [41 %] and n = 7 [19 %], respectively) for the whole cohort (p = 0.02), and for the gonadotroph cohort separately (n = 27 [42 %] and n = 6 [19 %], respectively, p = 0.03). There was no association between the gene expression of SOX2 and reintervention (p = 0.69). In the gonadotroph tumours, survival analyses showed that younger age (Exp(B) 0.95 [CI 0.92–0.98]) and absence of SOX2 staining (Exp(B) 0.35 [CI 0.14–0.89]), but not gender (Exp(B) 1.2 [CI 0.61-2.71]) were associated with a higher risk of reintervention (Fig. 5). We did not find an association between SOX9 or PROP1 and reintervention.
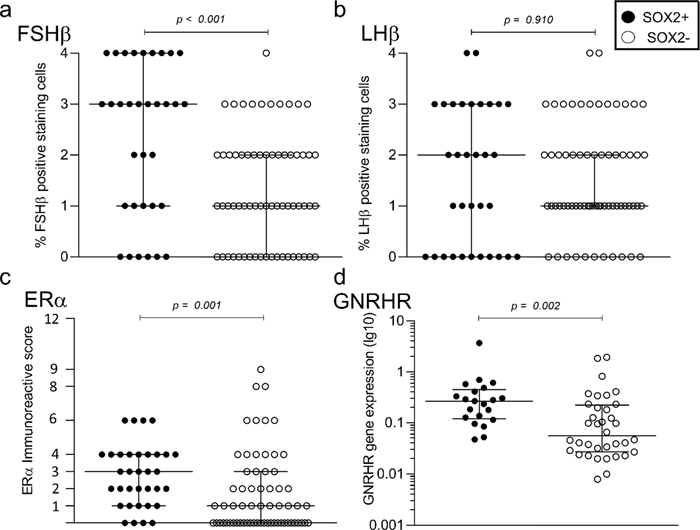
Figure 5: Kaplan-Meier curve showing the difference in reintervention rate between patients with gonadotroph tumours without SOX2 staining and tumours with SOX2 staining (p = 0.03). The number at risk at 25, 50, 100 and 150 months were 52, 40, 22 and eight, respectively, for the tumours not stained for SOX2, and 26, 24, 16 and three, respectively, for the tumours with SOX2 staining. Stem cell-associated markers and regulators of the gonadotroph axis For the gonadotroph NF-PitNETs, we found no difference in gender, age, tumour volume or invasiveness between tumours stained or not for the stem cell-associated markers SOX2, SOX9 or PROP1 (Suppl. Table 2). The staining for SOX2, SOX9 and PROP1 was associated with higher FSHβ but not LHβ gene expression and staining (Suppl. Table 2). SOX2-positive tumours showed higher levels of FSHβ and Estrogen Receptor α (ERα) staining as well as higher Estrogen Receptor 1 (ESR1) and GNRHR gene expression (Fig 6. a, c, d and Suppl. Table 2), whereas no difference was observed for LHβ (Fig 6 b). Similar patterns were observed for SOX9- and PROP1-positive tumours. No correlation was found between SOX2, SOX9, PROP1 and SF1 gene expression (Suppl. Table 2).
Figure 6: Expression of SOX2 and regulators of the gonadotroph axis. The presence of SOX2 was positively associated with higher levels of FSHβ, ERα and gene expression of GNRHR in the gonadotroph subgroup of NF-PitNETs (n = 101). SOX2 staining is classified as positive (staining score ≥ 1) or negative (staining score 0). Horizontal bars indicate median and interquartile values. Statistical comparisons are calculated by Mann-Whitney U test for all associations. Discussion The stem cell-associated markers SOX2, SOX9 and PROP1 were distinctly present, usually in a small proportion of cells, only in a subset of NF-PitNETs. The IHC staining and the gene expression levels correlated well for the same marker and between different markers. We did not find any association between the presence of the investigated markers, age at surgery, tumour volume or invasiveness. Absent SOX2 staining was associated with a higher rate of reintervention in the whole cohort of tumours and in the gonadotroph subgroup of tumours. The presence of the selected markers was positively associated with some of the regulators of the gonadotroph axis such as FSHβ, GNRHR and ERα in gonadotroph NF-PitNETs. In accordance with previous experimental studies [33–35], we demonstrate that cells with stem cell features are present at a low number in a subset of PitNETs. They tend to occur in small niches and have heterogeneous morphology corresponding to tumour cells and occasionally to endothelial cells. In the murine pituitary, SOX2-positive cells are present in a small proportion of cells (3–5 %) and usually scattered throughout the gland, and more concentrated lining the pituitary cleft [36]. A previous IHC study found SOX2 positivity in 20 % of human NF-PitNETs, which is not far from our results [19]. Interestingly, by using multiplex IHC, we demonstrate expression of the stem cell-associated markers SOX2 and SOX9 in intratumoural cells with epithelial features. However, in the cells positive for these markers, there was no immunolabeling for pituitary-specific transcription factors. It is not clear whether the cells coexpressing stem cell and epithelial markers may correspond to the previously described “follicular cells” of gonadotroph tumours, though these cells were found to express gonadotroph transcription factors in the study by Delfin et al [37]. Expression of stem cell-associated markers in a population of endothelial cells is in line with previous research, and may support speculation that transdifferentiation of stem cells into endothelial cells may contribute to tumour neovascularisation [33]. Moreover, SOX2 and SOX9 were strongly expressed in the epithelial cells of the Rathke's pouch remnants embedded in the tumour tissue, in accordance with previous studies demonstrating expression of stem cell markers in Rathke's pouch epithelium [18, 36] as well as in craniopharyngiomas originating from the epithelial remnants of Rathke's pouch [38]. The tumours lacking SOX2 staining presented a higher rate of reintervention in our cohort, contradictory to what we assumed initially. Surprisingly, Zhang et al found that the well differentiated SF1-tumours showed a higher rate of recurrence than the poorly differentiated tumours [18]. We have also previously presented data on markers associated with tumours being more differentiated and having a higher rate of reintervention in NF-PitNETs [30, 39]. The association between recurrence and differentiation in the SF1-tumours deviated from pituitary tumours of other cell lineages in the study from Zhang et al, corresponding to previous investigations in functioning pituitary tumours [40, 41]. Why the SF1-tumours deviate from the remaining pituitary tumours in this matter has not been explored. We found a moderate to strong correlation between the three markers SOX2, SOX9 and PROP1. Previous studies have shown that pituitary stem cells express both SOX2 and SOX9 [42], and cells with stem cell properties lining the borders of the marginal zone are also positive for PROP1 [1]. In our study, the presence of the stem cell-associated markers was consistently associated with markers of the gonadotroph cell phenotype. As previously described, stem cells lose SOX2 expression and generate committed progenitor cell lineages during embryonic development [43]. Recently it was hypothesised that SOX2-positive cells may give rise to a well organised structural and functional network that coordinates the responses to challenges, similar to the hormone secreting cells [5, 44]. Our data suggest that the SOX2-positive cell network is closely related to the gonadotroph cell network, or that both cell types respond to similar factors (e.g. nutrients, blood supply), and are located in similar areas in the parenchyma of the pituitary tumours. Whether the cells with stem cell features have a role in the hormonal regulation of the gonadotroph neoplastic cells, or whether the correlation presented here mirrors the normal pituitary phenotype is currently unknown. Limitations The study was of retrospective design, and tumour tissue was not available for all patients operated during the given time period, which may imply an inclusion bias. The IHC analyses were based on TMAs and not whole section slides. This might underestimate the presence of the markers investigated, knowing that stem cells or cells with stem cell features are few and might reside in niches in the tumour. However, the strong correlation between gene expression data and immunohistochemistry, as well as the high concordance between staining score in replicate TMA punches from the same tumour, as described in this study, supports the reliability of data acquired by TMA assessment. The expression of the proteins analysed by IHC was quantified based on different scales, i.e. ERα by immunoreactive score (IRS) and SOX2 by the percentage of distribution. This was due to a usually low number of positive cells and almost uniform strong staining, which makes IRS score unapplicable. Due to the low number of patients with radiological follow-up, we used the need for reintervention as the main end point concerning tumour aggressiveness. This might be influenced by the primary surgical result and by the operability of the patient. Tumour volume and invasiveness were not included in the regression analysis for the same reason. These variables would have been of great interest to be included in the analysis if they were available for the whole cohort. We did not succeed in performing IHC with endothelial markers as a part of multiplex IHC. Thus, we could not explore coexpression of endothelial and stem cell-associated markers in the endothelial-looking cells positive for stem cell markers. Conclusion The stem cell-associated markers SOX2, SOX9 and PROP1 are present at low levels in NF-PitNETs. They are correlated with each other, and their expression seems to be associated with the regulation of gonadotropins in the gonadotroph subgroup of tumours. Present or increased staining for the markers was not associated with tumour volume or invasiveness, though the absence of SOX2 seems to characterise tumours with a higher rate of reintervention. The stem cell-associated markers SOX2 and SOX9 are frequently coexpressed and are usually positive in the intratumoural cells with epithelial features lacking however immunolabeling for pituitary transcription factors. Declarations and statements Competing interests Olivera Casar-Borota is an associate editor of Free Neuropathology. The other authors declare no competing interests. Funding Olivera Casar-Borota was supported by the Swedish Cancer Society (grant number 190157), the Lions Cancerforskningsfond and a grant from the Swedish state under the agreement between the Swedish government and the county councils (ALF-agreement). For the remaining authors, no funding was received to assist with the preparation of this manuscript. Ethics approval This study was performed in line with the principles of the Declaration of Helsinki. Approval was granted by the regional Ethics committee for the south-east part of Norway (REK 2014/635), and by the hospital authorities. Consent to participate Informed consent was obtained from all living patients. Data sharing statement The data underlying this article cannot be shared publicly due to the privacy of individuals that participated in the study. The data will be shared on reasonable request to the corresponding author. Author contribution Kristin Astrid Øystese, Nicoleta Cristina Olarescu, Jens Petter Berg, Jens Bollerslev and Olivera Casar-Borota contributed to the study conception and design. Material preparation, data collection and analysis were performed by Kristin Astrid Øystese, Nicoleta Cristina Olarescu, Cecilia Lindskog, Fabjola Xheka, Jon Berg-Johnsen and Olivera Casar-Borota. The first draft of the manuscript was written by Kristin Astrid Øystese and all authors commented on previous versions of the manuscript. All authors read and approved the final manuscript. Acknowledgment M. Sc Jonas Gustavsson for performing multiplex IHC. References 1. Cox B, Roose H, Vennekens A, Vankelecom H. Pituitary stem cell regulation: who is pulling the strings? J Endocrinol. 2017;234(3):R135-r58. Epub 20170614. https://doi.org/10.1530/joe-17-0083. PubMed PMID: 28615294. 2. Garcia-Lavandeira M, Quereda V, Flores I, Saez C, Diaz-Rodriguez E, Japon MA, et al. A GRFa2/Prop1/stem (GPS) cell niche in the pituitary. PLoS One. 2009;4(3):e4815. Epub 20090313. https://doi.org/10.1371/journal.pone.0004815. PubMed PMID: 19283075; PubMed Central PMCID: PMC2654029. 3. Zhang S, Cui Y, Ma X, Yong J, Yan L, Yang M, et al. Single-cell transcriptomics identifies divergent developmental lineage trajectories during human pituitary development. Nat Commun. 2020;11(1):5275. Epub 20201019. https://doi.org/10.1038/s41467-020-19012-4. PubMed PMID: 33077725; PubMed Central PMCID: PMC7572359. 4. Ullah I, Subbarao RB, Rho GJ. Human mesenchymal stem cells - current trends and future prospective. Biosci Rep. 2015;35(2). Epub 20150428. https://doi.org/10.1042/bsr20150025. PubMed PMID: 25797907; PubMed Central PMCID: PMC4413017. 5. Le Tissier PR, Murray JF, Mollard P. A New Perspective on Regulation of Pituitary Plasticity: The Network of SOX2-Positive Cells May Coordinate Responses to Challenge. Endocrinology. 2022;163(8). https://doi.org/10.1210/endocr/bqac089. PubMed PMID: 35713880; PubMed Central PMCID: PMC9273012. 6. Vesper AH, Raetzman LT, Camper SA. Role of prophet of Pit1 (PROP1) in gonadotrope differentiation and puberty. Endocrinology. 2006;147(4):1654-63. Epub 20051229. https://doi.org/10.1210/en.2005-1080. PubMed PMID: 16384867. 7. Ward RD, Raetzman LT, Suh H, Stone BM, Nasonkin IO, Camper SA. Role of PROP1 in pituitary gland growth. Mol Endocrinol. 2005;19(3):698-710. Epub 20041209. https://doi.org/10.1210/me.2004-0341. PubMed PMID: 15591534. 8. Raetzman LT, Ward R, Camper SA. Lhx4 and Prop1 are required for cell survival and expansion of the pituitary primordia. Development. 2002;129(18):4229-39. https://doi.org/10.1242/dev.129.18.4229. PubMed PMID: 12183375. 9. Osamura RY, Grossman A, Korbonits M, Kovacs K, Lopes MB, Matsuno A, et al. Tumours of the pituitary gland. In: Lloyd RV, Osamura RY, Klöppel G, Rosai J, editors. WHO Classification of TUmours of Endocrine Organs. 4th ed. Lyon: International Agency for Research on Cancer (IARC); 2017. p. 11-63. 10. Tjornstrand A, Gunnarsson K, Evert M, Holmberg E, Ragnarsson O, Rosen T, et al. The incidence rate of pituitary adenomas in western Sweden for the period 2001-2011. Eur J Endocrinol. 2014;171(4):519-26. Epub 2014/08/03. https://doi.org/10.1530/eje-14-0144. PubMed PMID: 25084775. 11. Agustsson TT, Baldvinsdottir T, Jonasson JG, Olafsdottir E, Steinthorsdottir V, Sigurdsson G, et al. The epidemiology of pituitary adenomas in Iceland, 1955-2012: a nationwide population-based study. Eur J Endocrinol. 2015;173(5):655-64. Epub 2015/10/02. https://doi.org/10.1530/eje-15-0189. PubMed PMID: 26423473. 12. Nishioka H, Tamura K, Iida H, Kutsukake M, Endo A, Ikeda Y, et al. Co-expression of somatostatin receptor subtypes and estrogen receptor-alpha mRNAs by non-functioning pituitary adenomas in young patients. Mol Cell Endocrinol. 2011;331(1):73-8. Epub 2010/08/28. https://doi.org/10.1016/j.mce.2010.08.011. PubMed PMID: 20797424. 13. Sjostedt E, Bollerslev J, Mulder J, Lindskog C, Ponten F, Casar-Borota O. A specific antibody to detect transcription factor T-Pit: a reliable marker of corticotroph cell differentiation and a tool to improve the classification of pituitary neuroendocrine tumours. Acta Neuropathol. 2017. Epub 2017/08/22. https://doi.org/10.1007/s00401-017-1768-9. PubMed PMID: 28823042. 14. Scangas GA, Laws ER, Jr. Pituitary incidentalomas. Pituitary. 2013. Epub 2013/09/21. https://doi.org/10.1007/s11102-013-0517-x. PubMed PMID: 24052242. 15. Melmed S. Pituitary-Tumor Endocrinopathies. N Engl J Med. 2020;382(10):937-50. Epub 2020/03/05. https://doi.org/10.1056/NEJMra1810772. PubMed PMID: 32130815. 16. Guido CB, Sosa LDV, Perez PA, Zlocoswki N, Velazquez FN, Gutierrez S, et al. Changes of stem cell niche during experimental pituitary tumor development. J Neuroendocrinol. 2021;33(12):e13051. Epub 20211027. https://doi.org/10.1111/jne.13051. PubMed PMID: 34708474. 17. Yunoue S, Arita K, Kawano H, Uchida H, Tokimura H, Hirano H. Identification of CD133+ cells in pituitary adenomas. Neuroendocrinology. 2011;94(4):302-12. Epub 20110908. https://doi.org/10.1159/000330625. PubMed PMID: 21912092. 18. Zhang Q, Yao B, Long X, Chen Z, He M, Wu Y, et al. Single-cell sequencing identifies differentiation-related markers for molecular classification and recurrence prediction of PitNET. Cell Rep Med. 2023;4(2):100934. Epub 20230207. https://doi.org/10.1016/j.xcrm.2023.100934. PubMed PMID: 36754052; PubMed Central PMCID: PMC9975294. 19. Capatina C, Cimpean AM, Raica M, Coculescu M, Poiana C. SOX 2 Expression in Human Pituitary Adenomas-Correlations With Pituitary Function. In Vivo. 2019;33(1):79-83. https://doi.org/10.21873/invivo.11442. PubMed PMID: 30587606; PubMed Central PMCID: PMC6364058. 20. Peverelli E, Giardino E, Treppiedi D, Meregalli M, Belicchi M, Vaira V, et al. Dopamine receptor type 2 (DRD2) and somatostatin receptor type 2 (SSTR2) agonists are effective in inhibiting proliferation of progenitor/stem-like cells isolated from nonfunctioning pituitary tumors. Int J Cancer. 2017;140(8):1870-80. Epub 2017/01/26. https://doi.org/10.1002/ijc.30613. PubMed PMID: 28120505. 21. Moncho-Amor V, Chakravarty P, Galichet C, Matheu A, Lovell-Badge R, Rizzoti K. SOX2 is required independently in both stem and differentiated cells for pituitary tumorigenesis in p27-null mice. Proc Natl Acad Sci U S A. 2021;118(7). https://doi.org/10.1073/pnas.2017115118. PubMed PMID: 33574062; PubMed Central PMCID: PMC7896314. 22. Nys C, Lee YL, Roose H, Mertens F, De Pauw E, Kobayashi H, et al. Exploring stem cell biology in pituitary tumors and derived organoids. Endocr Relat Cancer. 2022;29(7):427-50. Epub 20220617. https://doi.org/10.1530/erc-21-0374. PubMed PMID: 35521774. 23. Oystese KA, Zucknick M, Casar-Borota O, Ringstad G, Bollerslev J. Early postoperative growth in non-functioning pituitary adenomas; A tool to tailor safe follow-up. Endocrine. 2017;57(1):35-45. Epub 2017/05/19. https://doi.org/10.1007/s12020-017-1314-5. PubMed PMID: 28516382. 24. Knosp E, Steiner E, Kitz K, Matula C. Pituitary adenomas with invasion of the cavernous sinus space: a magnetic resonance imaging classification compared with surgical findings. Neurosurgery. 1993;33(4):610-7; discussion 7-8. Epub 1993/10/01. PubMed PMID: 8232800. 25. Micko AS, Wohrer A, Wolfsberger S, Knosp E. Invasion of the cavernous sinus space in pituitary adenomas: endoscopic verification and its correlation with an MRI-based classification. J Neurosurg. 2015;122(4):803-11. Epub 2015/02/07. https://doi.org/10.3171/2014.12.jns141083. PubMed PMID: 25658782. 26. Casar-Borota O, Fougner SL, Bollerslev J, Nesland JM. KIT protein expression and mutational status of KIT gene in pituitary adenomas. Virchows Arch. 2012;460(2):171-81. Epub 2012/01/04. https://doi.org/10.1007/s00428-011-1185-8. PubMed PMID: 22213031. 27. Kampf C, Olsson I, Ryberg U, Sjöstedt E, Pontén F. Production of tissue microarrays, immunohistochemistry staining and digitalization within the human protein atlas. J Vis Exp. 2012;(63). Epub 20120531. https://doi.org/10.3791/3620. PubMed PMID: 22688270; PubMed Central PMCID: PMC3468196. 28. Uhlén M, Fagerberg L, Hallström BM, Lindskog C, Oksvold P, Mardinoglu A, et al. Proteomics. Tissue-based map of the human proteome. Science. 2015;347(6220):1260419. https://doi.org/10.1126/science.1260419. PubMed PMID: 25613900. 29. Oystese KA, Casar-Borota O, Normann KR, Zucknick M, Berg JP, Bollerslev J. Estrogen Receptor alpha, a Sex-Dependent Predictor of Aggressiveness in Nonfunctioning Pituitary Adenomas: SSTR and Sex Hormone Receptor Distribution in NFPA. J Clin Endocrinol Metab. 2017;102(9):3581-90. Epub 2017/09/16. https://doi.org/10.1210/jc.2017-00792. PubMed PMID: 28911153. 30. Oystese KAB, Berg JP, Normann KR, Zucknick M, Casar-Borota O, Bollerslev J. The role of E and N-cadherin in the postoperative course of gonadotroph pituitary tumours. Endocrine. 2018;62(2):351-60. Epub 2018/07/28. https://doi.org/10.1007/s12020-018-1679-0. PubMed PMID: 30051197. 31. Bustin SA, Benes V, Garson JA, Hellemans J, Huggett J, Kubista M, et al. The MIQE guidelines: minimum information for publication of quantitative real-time PCR experiments. Clin Chem. 2009;55(4):611-22. Epub 2009/02/28. https://doi.org/10.1373/clinchem.2008.112797. PubMed PMID: 19246619. 32. Normann KR, Oystese KA, Berg JP, Lekva T, Berg-Johnsen J, Bollerslev J, et al. Selection and validation of reliable reference genes for RT-qPCR analysis in a large cohort of pituitary adenomas. Mol Cell Endocrinol. 2016;437:183-9. Epub 2016/08/27. https://doi.org/10.1016/j.mce.2016.08.030. PubMed PMID: 27561203. 33. Mertens F, Gremeaux L, Chen J, Fu Q, Willems C, Roose H, et al. Pituitary tumors contain a side population with tumor stem cell-associated characteristics. Endocr Relat Cancer. 2015;22(4):481-504. Epub 20150428. https://doi.org/10.1530/erc-14-0546. PubMed PMID: 25921430. 34. Chen J, Gremeaux L, Fu Q, Liekens D, Van Laere S, Vankelecom H. Pituitary progenitor cells tracked down by side population dissection. Stem Cells. 2009;27(5):1182-95. https://doi.org/10.1002/stem.51. PubMed PMID: 19418455. 35. Carreno G, Gonzalez-Meljem JM, Haston S, Martinez-Barbera JP. Stem cells and their role in pituitary tumorigenesis. Mol Cell Endocrinol. 2017;445:27-34. Epub 20161006. https://doi.org/10.1016/j.mce.2016.10.005. PubMed PMID: 27720895. 36. Fauquier T, Rizzoti K, Dattani M, Lovell-Badge R, Robinson IC. SOX2-expressing progenitor cells generate all of the major cell types in the adult mouse pituitary gland. Proc Natl Acad Sci U S A. 2008;105(8):2907-12. Epub 20080215. https://doi.org/10.1073/pnas.0707886105. PubMed PMID: 18287078; PubMed Central PMCID: PMC2268558. 37. Delfin L, Mete O, Asa SL. Follicular cells in pituitary neuroendocrine tumors. Hum Pathol. 2021;114:1-8. Epub 2021/05/16. https://doi.org/10.1016/j.humpath.2021.05.002. PubMed PMID: 33991528. 38. Garcia-Lavandeira M, Saez C, Diaz-Rodriguez E, Perez-Romero S, Senra A, Dieguez C, et al. Craniopharyngiomas express embryonic stem cell markers (SOX2, OCT4, KLF4, and SOX9) as pituitary stem cells but do not coexpress RET/GFRA3 receptors. J Clin Endocrinol Metab. 2012;97(1):E80-7. Epub 20111026. https://doi.org/10.1210/jc.2011-2187. PubMed PMID: 22031517. 39. Øystese KAB, Casar-Borota O, Berg-Johnsen J, Berg JP, Bollerslev J. Distribution of E- and N-cadherin in subgroups of non-functioning pituitary neuroendocrine tumours. Endocrine. 2022;77(1):151-9. Epub 2022/06/09. https://doi.org/10.1007/s12020-022-03051-6. PubMed PMID: 35674926; PubMed Central PMCID: PMC9242907. 40. Evang JA, Berg JP, Casar-Borota O, Lekva T, Kringen MK, Ramm-Pettersen J, et al. Reduced levels of E-cadherin correlate with progression of corticotroph pituitary tumours. Clin Endocrinol (Oxf). 2011;75(6):811-8. Epub 2011/05/21. https://doi.org/10.1111/j.1365-2265.2011.04109.x. PubMed PMID: 21595730. 41. Fougner SL, Lekva T, Borota OC, Hald JK, Bollerslev J, Berg JP. The expression of E-cadherin in somatotroph pituitary adenomas is related to tumor size, invasiveness, and somatostatin analog response. J Clin Endocrinol Metab. 2010;95(5):2334-42. Epub 2010/03/26. https://doi.org/10.1210/jc.2009-2197. PubMed PMID: 20335450. 42. Zhang Z, Zamojski M, Smith GR, Willis TL, Yianni V, Mendelev N, et al. Single nucleus transcriptome and chromatin accessibility of postmortem human pituitaries reveal diverse stem cell regulatory mechanisms. Cell Rep. 2022;38(10):110467. Epub 2022/03/10. https://doi.org/10.1016/j.celrep.2022.110467. PubMed PMID: 35263594; PubMed Central PMCID: PMC8957708. 43. Russell JP, Lim X, Santambrogio A, Yianni V, Kemkem Y, Wang B, et al. Pituitary stem cells produce paracrine WNT signals to control the expansion of their descendant progenitor cells. Elife. 2021;10. Epub 20210105. https://doi.org/10.7554/eLife.59142. PubMed PMID: 33399538; PubMed Central PMCID: PMC7803373. 44. Mollard P, Hodson DJ, Lafont C, Rizzoti K, Drouin J. A tridimensional view of pituitary development and function. Trends Endocrinol Metab. 2012;23(6):261-9. Epub 20120320. https://doi.org/10.1016/j.tem.2012.02.004. PubMed PMID: 22436593.
Copyright: © 2024 The author(s). This is an open access article distributed under the terms of the Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited, a link to the Creative Commons license is provided, and any changes are indicated. The Creative Commons Public Domain Dedication waiver (https://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||