|
|
|||||||||||||||||||||||||||||||||||||
|
Free Neuropathology 5:10 (2024) |
|||||||||||||||||||||||||||||||||||||
|
Review |
|||||||||||||||||||||||||||||||||||||
|
Fluid preservation in brain banking: a review |
|||||||||||||||||||||||||||||||||||||
|
Andrew T. McKenzie1, Oge Nnadi2, Kat D. Slagell3,4,5, Emma L. Thorn3,4,5, Kurt Farrell3,4,5, John F. Crary3,4,5 |
|||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||
|
Corresponding author: |
|||||||||||||||||||||||||||||||||||||
|
Additional resources and electronic supplementary material: supplementary material |
|||||||||||||||||||||||||||||||||||||
|
Submitted: 12 February 2024 |
|||||||||||||||||||||||||||||||||||||
|
Keywords: Biobanking, Postmortem brain, Fluid preservation, Formaldehyde, Overfixation, Storage artifact, Tissue clearing, Glycerol |
|||||||||||||||||||||||||||||||||||||
|
Abstract Fluid preservation is nearly universally used in brain banking to store fixed tissue specimens for future research applications. However, the effects of long-term immersion on neural circuitry and biomolecules are not well characterized. As a result, there is a need to synthesize studies investigating fluid preservation of brain tissue. We searched PubMed and other databases to identify studies measuring the effects of fluid preservation in nervous system tissue. We categorized studies based on the fluid preservative used: formaldehyde solutions, buffer solutions, alcohol solutions, storage after tissue clearing, and cryoprotectant solutions. We identified 91 studies containing 197 independent observations of the effects of long-term storage on cellular morphology. Most studies did not report any significant alterations due to long-term storage. When present, the most frequent alteration was decreased antigenicity, commonly attributed to progressive crosslinking by aldehydes that renders biomolecules increasingly inaccessible over time. To build a mechanistic understanding, we discuss biochemical aspects of long-term fluid preservation. A subset of lipids appears to be chemical altered or extracted over time due to incomplete retention in the crosslinked gel. Alternative storage fluids mitigate the problem of antigen masking but have not been extensively characterized and may have other downsides. We also compare fluid preservation to cryopreservation, paraffin embedding, and resin embedding. Overall, existing evidence suggests that fluid preservation provides maintenance of neural architecture for decades, including precise structural details. However, to avoid the well-established problem of overfixation caused by storage in high concentration formaldehyde solutions, fluid preservation procedures can use an initial fixation step followed by an alternative long-term storage fluid. Further research is warranted on optimizing protocols and characterizing the generalizability of the storage artifacts that have been identified. |
|||||||||||||||||||||||||||||||||||||
|
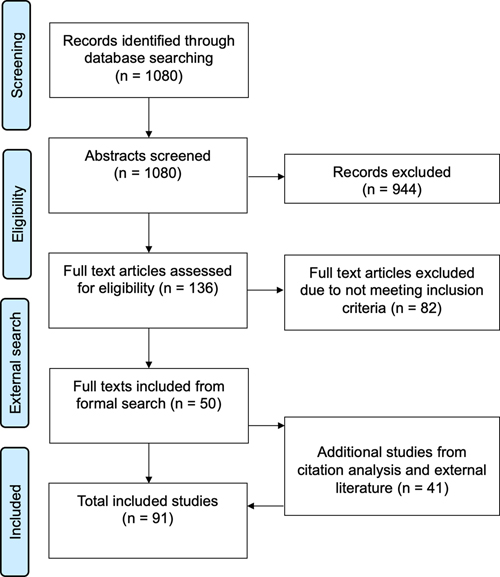
Introduction Fluid preservation, or storage in liquid, is a common method of preserving biological specimens intended for research and education. In global natural history collections, a substantial percentage of the billions of specimens are maintained in a fluid state, often referred to as “wet collections” (Marte et al., 2003; Hilton et al., 2021). In brain banking, fluid preservation stands alongside paraffin embedding and cryopreservation as one of the major preservation methods. Fluid preservation is commonly used because it is simple to perform, inexpensive, and suitable for many investigations. Worldwide, there are many thousands of brains preserved in fluid. For example, the University of Geneva collection contains over 10,000 human brains preserved in formalin (Kövari et al., 2011). Despite the ubiquity of fluid preservation, no one method for preserving brains in the fluid state has yet established itself as obviously the best choice. Instead, many different methods are used, each with advantages and disadvantages. The long-term storage outcomes of these fluid preservation methods in comparison with each other and with non-fluid state storage methods remain unclear. Therefore, there is a critical need to comprehensively investigate the effect of fluid preservation methods in the long-term storage of brain tissue, including their impact on preserving cellular and molecular structures, subsequent data analysis, and the accumulation of any storage artifacts over time. In this review, our focus is the fluid preservation of fixed brains, drawing upon a distinction between initial fixation methods and long-term storage or preservation methods outlined in the biobanking literature (Hartman, 2019). “Fixation” is used in the specific sense of covalent crosslinking of biomolecules with a fixative such as formaldehyde (Stoddart, 1989). This initial fixation process, which can involve immersion or perfusion, is largely independent of the long-term storage method (McFadden et al., 2019; McKenzie et al., 2022). A practical working definition dividing initial fixation from long-term preservation is the period after which the tissue can be removed from the fixative but remain intact. However, there is often a grey area between the initial fixation and long-term storage. For example, brains are sometimes left in the same fixative medium for long-term storage. Furthermore, there may be no clear boundary when fixation is “complete”, as fixation strength is on a spectrum without clear thresholds. To understand a complex field such as fluid preservation, it is helpful to review the evolution of the methods (Carlos et al., 2019). Frederik Ruysch, a Dutch anatomist and pioneer in biospecimen preservation, was among the first to use a fluid containing alcohol for preservation during the 17th century (Luyendijk-Elshout, 1970). Alcohol was the preferred preservative until the 1890s, when the fixative effects of the newly commercially available formaldehyde were discovered by Ferdinand Blum (Baird, 1859; Fox et al., 1985; Herbin et al., 2021). Formaldehyde was preferred because it was less expensive, non-flammable, and led to better preservation of morphology, so its use rapidly expanded (Fish, 1895). Formaldehyde also began to be used for long-term fluid preservation (Herbin et al., 2021). The choice of ideal preservation fluid for long-term brain banking has not received as much attention in recent years, but this is worthy of re-evaluation given the increased emphasis on reproducibility in modern research. The ideal method involves striking a balance between various goals: safeguarding the specimen's biomolecular structure, maintaining chemical stability, and ensuring safe handling via low flammability, toxicity, and volatility. It should also be cost-effective, user-friendly, and compatible with downstream applications, which may not always be pre-defined in prospective biobanking projects. Novelty is also considered a downside for preservation methods, as long-term preservation outcomes are best evaluated with time. Several candidate chemicals or chemical combinations have shown promise in meeting these criteria, each with their own advantages and drawbacks. In this review, we aim to dissect the current body of knowledge surrounding fluid preservation techniques for brain specimens, with an emphasis on the maintenance of morphomolecular characteristics for brain mapping studies. In recent years, several next-generation brain mapping technologies have emerged, allowing for the 3D visualization of the cellular and molecular organization of the brain (Shapson-Coe et al., 2021; Patel et al., 2022). Although existing reviews have focused on the long-term fluid preservation of biomolecules such as DNA, it is less clear how fluid preservation in brain banking will affect the neural structures visualized with these cutting-edge brain mapping methods (Lou et al., 2014; Gustafsson et al., 2015). Accurate and reliable mapping requires specimens that have been preserved in a manner that maintains their original cellular and molecular structures as closely as possible. This review may be useful to professionals engaged in the development and management of brain collections, as it offers guidance in selecting preservation techniques. Moreover, researchers studying tissue from existing brain banks, particularly those with an interest in unraveling neural circuitry from archival brain samples, may find this review helpful in guiding experimental design. This review is aimed to help ensure the reliability of data generated from these invaluable resources, ultimately supporting the development of more effective therapies for neurobiological disorders. Methods and literature search We conducted a realist synthesis review with the goal of developing a theoretical understanding of fluid preservation, an approach that combines aspects of a systematic review with a focus on theory and applicability (Wong et al., 2013). This style was selected given the broad and variably defined nature of fluid preservation. We followed the RAMESES reporting standards (see Supplementary File 1) (Wong et al., 2013). Before formalizing our search methodology, we first scoped the literature by searching PubMed, Google Scholar, bioRxiv, and medRxiv, and by holding discussions among the authors. The review protocol was preregistered here: https://osf.io/jdyvs. Additional review methods are available in Supplementary File 2. Through our formal search process, we screened 1080 abstracts, reviewed 136 full texts, and included 50 studies (Figure 1). We also identified 41 studies through citation analysis or ad hoc searches, after which we eventually included 91 total studies (Supplementary Files 3-6). We categorized the included studies into five types of chemicals used for storage: formaldehyde-containing solutions (n=68), buffer (n=5), alcohol-containing solutions (n=3), storage after tissue clearing (n=5), and cryoprotectant-containing solutions (n=10). Collectively, these studies contain 197 distinct observations about the effects of fluid preservation on cellular structure. Prior to discussing outcomes for each of these storage options, we first review some relevant biochemistry.
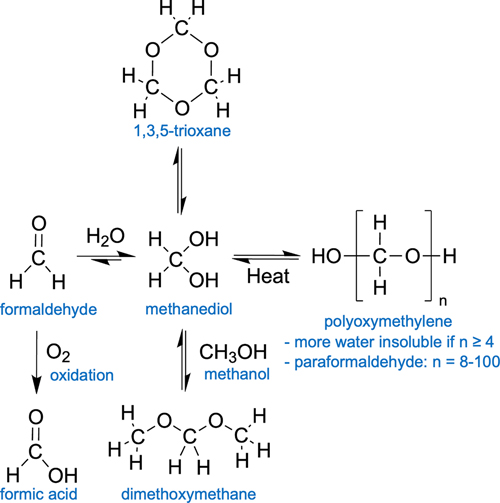
Figure 1. Study selection flow diagram. Biochemistry of long-term fluid preservation Polymer properties of formaldehyde Formaldehyde is by far the most common initial preservative used in brain banking. Molecular formaldehyde is a gas that dissolves rapidly in water. In water, it is rapidly hydrated to form methanediol, also known as methylene glycol or formaldehyde monohydrate (Figure 2). In aqueous solutions, methanediol is abundant, while unhydrated monomeric formaldehyde is almost completely absent (Boyer et al., 2013; Walker, 1944). In turn, methanediol oligomerizes or polymerizes very rapidly, making it difficult to isolate in the pure state (Schmitz et al., 2015). Specifically, methanediol reacts to form 1,3,5-trioxane, a stable cyclic trimer, or polyoxymethylene glycol, which is a polymer form with a variable number of subunits. The equilibrium of monomeric and polymeric formaldehyde hydrates in a solution is governed by factors such as the temperature and concentration of formaldehyde. At higher temperature and more dilute concentrations, methanediol is favored, thus leading to the depolymerization of polyoxymethylene glycol. Additionally, in an alkaline environment, hydroxyl end-groups of the polymer form are more rapidly degraded, leading to the progressive cleavage of formaldehyde units from the extremities of the linear molecular chains (French and Edsall, 1945).
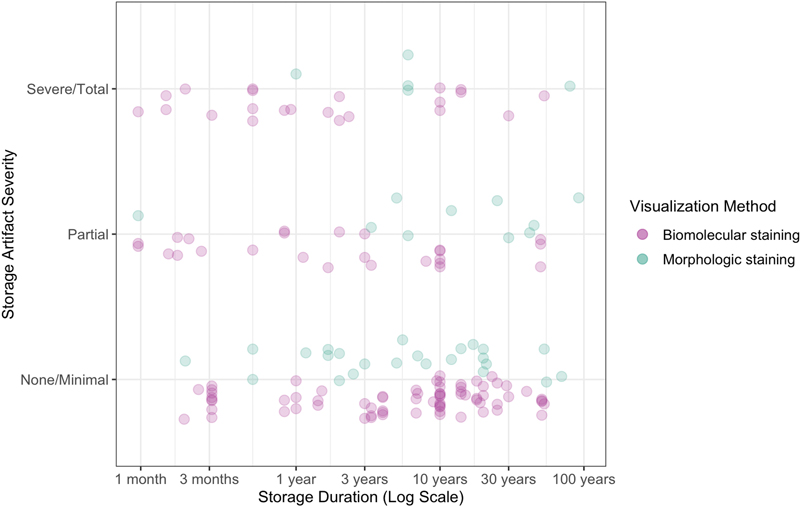
Figure 2. Schematic diagram of chemical alterations of formaldehyde in aqueous solutions. Paraformaldehyde is a commercial storage form of polyoxymethylene, with a mixture of polymerization products with degrees of 8-100 formaldehyde units (Walker, 1944). It is thought to be the monomeric form that is reactive and crosslinks biomolecules. As a result, the polyoxymethylene polymers in paraformaldehyde must be depolymerized prior to use as a fixative. This usually involves heating and the addition of sodium hydroxide to increase the pH of the solution and stimulate cleavage of formaldehyde units. When longer polyoxymethylene polymers form in aqueous solutions of formaldehyde, which is more likely to occur during storage at low temperatures, they can become insoluble and precipitate out of the solution. Methanol is often added to formaldehyde as a stabilizer. For example, the commercial product formalin often contains 37 % formaldehyde by weight and approximately 10 % of methanol. Methanol is believed to prevent polymer precipitation in formaldehyde solutions through the reaction of the alcohol group in methanol with the aldehyde group in formaldehyde, leading to the formation of hemiacetals or acetals such as dimethoxymethane (Norris et al., 2010; Walker, 1944). These hemiacetals interfere with the polymerization process of methanediol, thereby stabilizing them in solution and preventing the formation and precipitation of solid polymers. Notably, other chemicals that also contain an alcohol group, such as ethanol, glycols, and glycerol, can also stabilize formaldehyde and limit the extent of its polymerization. The reason that methanol is most commonly used as a stabilizer appears to be partially a historical consequence of the fact that formaldehyde was manufactured from methanol (Walker, 1944). A complicating factor is that methanol itself acts a fixative and may also contribute to tissue decomposition over time, especially of lipids. Some investigators use depolymerized paraformaldehyde that lacks any methanol. The practical effect of the methanol stabilizer in fixation and long-term storage of biospecimens is not well established. Formic acid In addition to polymerization, formaldehyde solutions stored at room temperature can undergo several chemical other reactions, leading to the formation of methanol, hydroxyaldehydes, sugars, and formic acid (Walker, 1944). For our purposes, the most important chemical reaction is thought to be the formation of formic acid. This can occur through two pathways. First, formaldehyde can be oxidized via oxygen in the solution to formic acid. Second, even if substantial oxygen is not present, the Cannizzaro reaction can occur, which is a disproportionation reaction wherein one molecule of formaldehyde is oxidized to formic acid, and another is reduced to methanol. Formic acid formation is a problem for brains stored in formaldehyde solutions over the long-term for several reasons. First, formic acid has the potential to solubilize proteins, especially hydrophobic ones, such as those found in myelin (Zheng and Doucette, 2016). Second, formic acid can also induce chemical modifications to proteins, which limits the use of formic acid in proteomics (Zheng and Doucette, 2016). As an example of this, formic acid has been found to destroy prion proteins in a way that formaldehyde itself does not (Taylor et al., 1997). Although formic acid may not adversely affect histology after short exposure times of one hour, its long-term effects on brain tissue preservation are still a concern (Brown et al., 1990). One source notes that the long-term storage of biospecimens in formaldehyde is corrosive because it breaks down into formic acid (Eichhorn et al., 2018). One study used a novel formulation of formaldehyde that removed formic acid with an ion-exchange basic resin, which they described as acid-deprived formaldehyde (Berrino et al., 2022). In this study, after six months of storage, they reported significantly less DNA fragmentation in the samples stored in formic acid-deprived formaldehyde compared to standard formaldehyde solutions. Notably, RNA was stable over the six-month period tested when stored in either solution. pH buffering In addition to directly damaging tissue, the formation of formic acid over time will also lower the pH of a solution containing formaldehyde. In turn, lower pH can damage tissue by causing or accelerating protein denaturation, protein aggregation, and the hydrolysis of biomolecules. As a result, it is commonly thought that more acidic conditions are associated with worse tissue degradation (van Duijn et al., 2011). To prevent acidification of formaldehyde solutions, a neutral buffer is often used, which is often in the form of a phosphate buffer. A pH buffer system involves the use of weak acids and their corresponding conjugate bases, or vice versa. In the case of phosphate buffer, monohydrogen phosphate, [PO3(OH)]2-, acts as the conjugate acid and dihydrogen phosphate, [H2PO4]-, acts as the conjugate base. These ions form a buffer system that can slow changes in pH. Formalin with a phosphate buffer added is called neutral buffered formalin (NBF). Unbuffered formalin has a pH of approximately 3.7, while that of buffered formalin can be tuned, usually to a pH of approximately 7.0 (Nuovo and Richart, 1989). Historically, NBF was not widely used, but it has been more commonly used in the past several decades. NBF is expected to lead to less tissue damage during storage by preventing a drop in pH as the formaldehyde-preserved solution forms acids. However, the capacity of the buffer system will eventually be reached, and with additional acid formation, the pH will decrease regardless of the initial buffer. Multiple empirical studies have examined the acidification of formalin solutions, both buffered and unbuffered, in the preservation of human brains. These studies have found that the pH of brain tissue fixed with buffered formalin tends to decrease over time. For example, one study found that the pH of buffered formalin solutions was 6.4 after 1 month, 5.7 at 8 years, and 4.5 after 10 years (Pikkarainen et al., 2010). Another study, profiling brains with fixation times of 7 months to more than 50 years, found that the pH of the formalin solutions had a pH range of 5 in the more recently preserved brains to 4 in the older cases (Sheaffer et al., 1999). Other studies have reported similar pH ranges from 4-6 that decrease over time (Ploeger et al., 1993; Rosoklija et al., 2003). Factors contributing to the differences in the rate of pH decline across collections likely include the volume and concentration of fixative, the type of brain tissue, the container used, and the amount of oxygen exposure. Notably, one source notes that this drop of pH in stored formalin solutions only occurs in the presence of tissue (Weil, 1929). This finding suggests that there is an interaction between formaldehyde solutions and tissue biomolecules that promotes acid formation. Taken together, the use of phosphate buffer can slow, but not prevent, the lowering of pH in formalin solutions when they are used to preserve brains for the long-term. Initial aldehyde crosslinking of biomolecules The initial fixation step in brain banking is typically performed using formaldehyde, which is favored over alternative aldehyde fixatives due to its high diffusion rate. Formaldehyde covalently crosslinks intracellular and extracellular biomolecules, forming both intramolecular and intermolecular bonds (Walker, 1944). The exact chemical mechanisms remain somewhat uncertain and context-dependent, but some principles can be identified (Gustafsson et al., 2015; Tayri-Wilk et al., 2020). Formaldehyde predominantly crosslinks arginine and lysine side chains of proteins, followed by tyrosine and aspartic acid (Tayri-Wilk et al., 2020). Although methylene bridges between two amino acids (R1-CH2-R2) were previously believed to be the primary crosslinks, recent studies suggest a higher prevalence of formaldehyde reacting to form imine bonds (R-N=CH2) on distinct amino acids, which then crosslink through an unknown mechanism (Tayri-Wilk et al., 2020). The ability of formaldehyde to react with the amino and imino groups of DNA also enables it to crosslink DNA with proteins, making it integral to chromatin studies (Hoffman et al., 2015). For example, there is minimal free DNA (<10 %) detected after just minutes of formaldehyde fixation (as cited in Hoffman et al., 2015). Prolonged or concentrated exposure to formaldehyde leads to the formation of insoluble higher-order crosslinked chromatin complexes (Hoffman et al., 2015; Nilsen, 2014). The effects of initial fixation on different lipid species in brain tissue is variable. Phosphatidylethanolamines, and to a lesser extent phosphatidylserines, which contain primary amines, readily undergo crosslinking with proteins and other biomolecules through their amine groups (Carter et al., 2016; Vos et al., 2019; Bien et al., 2021; Kotnala et al., 2021). These lipids likely form an interconnected mesh with other cross-linked proteins, making them challenging to detect in their free form after fixation using approaches like mass spectrometry that require dissociated molecules, in the absence of antigen retrieval steps (Carter et al., 2016; Denti et al., 2020). In contrast, most other lipid types lacking amines, such as phosphatidylcholine, sphingomyelin, and cholesterol, have minimal direct crosslinking upon initial fixation, as suggested by the minimal changes in their abundance on mass spectrometry or magnetic resonance after fixation (Purea and Webb, 2006; Bien et al., 2021; Kotnala et al., 2021). However, some of these lipids may be indirectly retained in the tissue through the protein-amine phospholipid-crosslink mesh, though this depends on the harshness of subsequent processing steps (Maneta-Peyret et al., 1999). In addition, initial fixation does not cause significant changes in the spatial localization of lipids, though species that have more membrane fluidity, such as cholesterol, can migrate during subsequent processing steps (Carter et al., 2011; Vos et al., 2019; Bien et al., 2021). Chemical alterations like hydrolysis, peroxidation, and methylation of amine head groups also likely occur for some lipids during initial fixation (Carter et al., 2016; Kotnala et al., 2021; Dannhorn et al., 2022). Chemical gel formation Because fixatives induce gel formation in tissues, the properties of gels are essential to understand in fluid preservation. A gel is a material primarily composed of liquid – often accounting for over 99 % of its total mass – constrained by a three-dimensional immobilizing matrix (Adams, 2022). The matrix bestows upon gels its key solid-like characteristic of elasticity, which enables the gel to resume its original form after deformation (Clark, 1991). Gel robustness or strength can be evaluated based on the extent of its structural deformation in response to mechanical stressors, such as inversion (Jia et al., 2011). Gels can be classified into chemical and physical types. Chemical gels originate from covalent crosslinks between chains, while physical gels originate from non-covalent interactions, such as electrostatic or hydrophobic forces (Gulrez et al., 2011; Richtering and Saunders, 2014). The crosslinks in chemical gels disintegrate at a significantly slower pace than in their physical counterparts, owing to the tough nature of covalent bonds. As a result, the covalent bonds in chemical gels are sometimes considered "essentially permanent" (Adams, 2022). On a physical level, gel formation triggers a steep increase in viscosity and effectively halts molecular motion among the crosslinked molecules, mirroring other liquid-solid phase transitions like colloidal aggregation or the glass transition (Trappe et al., 2001). Even prior to fixation, cells and extracellular matrix in biological tissues tend to have gel-like properties, which is not surprising because gels are the quintessential form of soft matter (Douglas, 2018). For example, the cytoskeleton of dendritic spines has been found to behave as a gel (Eberhardt et al., 2022). Following crosslinking fixation, these native gel-like networks are dramatically strengthened and stabilized (Wang and Minassian, 1987). As an example of this, crosslinking of chitosan with glutaraldehyde induces the formation of a chemical gel that strengthens with increased concentration of glutaraldehyde and time (Argüelles-Monal et al., 1998). An unfixed and sufficiently decomposed brain does not, in practice, act as a chemical gel. However, following the covalent crosslinking of proteins in the brain with formaldehyde, a fixed brain could be considered to have formed a chemical gel network. Indeed, once the brain has been fixed with formaldehyde and delipidated, it has been found to act macroscopically as a hydrogel, insofar as it can reversibly swell and shrink in water (Susaki et al., 2020). For a chemical gel in general, the number of covalent crosslinks, the distance between them, and its resulting strength, can be controlled by the chemistry used to create the gel matrix (Adams, 2022). Therefore, the crosslinking properties and the strength of the gel created by fixation depends on the aldehyde fixative(s) used and the duration of fixation. There are many trade-offs involved in determining the optimal duration, and thus the strength, of fixation. Longer fixation times are associated with more retention of antigens in the tissue prior to long-term storage or subsequent tissue processing steps (Romijn et al., 1999). On the other hand, longer fixation times are also associated with substantially decreased antibody penetration and reduced antigenicity for immunohistochemistry, which is a phenomenon called “overfixation” (Romijn et al., 1999; McFadden et al., 2019). In practice, 1-2 weeks of formaldehyde immersion is often considered necessary for “complete fixation” of the human brain at room temperature (Romijn et al., 1999). One group tested various fixation times ranging from 3 days to several months, finding that 6-14 days of fixation was the best duration of fixation for their goals (Romijn et al., 1999). Taken together, the duration of initial fixation is an important variable to consider in evaluating the literature on long-term fluid preservation. Chemical gel-based preservation of biomolecules over time We can consider two features of biomolecules that can be preserved: their chemical and location properties. Chemical refers to whether the atomic composition or conformation of a molecule has been modified during the fluid preservation process. This is relevant in fluid preservation, because in the liquid state, chemical reactions will still be occurring. Location refers to the relative position of a biomolecule in the tissue. Changes in biomolecular location during storage of fixed brains are dependent on the extent to which the biomolecule is incorporated into the crosslinked mesh. A pure chemical gel typically contains two components: an immobile network of crosslinked molecules and a mobile solvent. In contrast, we can categorize biomolecules in the fixed brain into three groups: (a) an immobile network of directly crosslinked biomolecules, which primarily consists of proteins, (b) a mobile solvent, which is usually water, but this can be replaced, and (c) entangled biomolecules that are not directly crosslinked but are indirectly confined in the protein mesh. This entangled category includes many lipids. If a protein is directly crosslinked, then the main question for maintenance of its location is the stability of its covalent bonds. In aqueous environments, the primary mechanism for the degradation of peptide bonds and crosslinking bonds is expected to be hydrolysis (Shiurba et al., 1998). Uncatalyzed peptide bond cleavage occurs at a slow rate; at neutral pH, hydrolysis of peptide bonds has demonstrated half-lives on the order of hundreds of years (Mahesh et al., 2018; Radzicka and Wolfenden, 1996). Enzymatic catalysis, which theoretically could accelerate the process, is anticipated to be substantially impeded by the crosslinking procedure. As a result, there is a strong theoretical rationale to expect that proteins exhibit considerable stability during fluid preservation. Hydrolysis of crosslinking bonds can certainly occur, especially at higher temperatures (Barker et al., 1980; Hoffman et al., 2015). However, in many contexts, a subset of aldehyde crosslinks are also expected to last for a substantial period (Shiurba et al., 1998). This is difficult to measure when the brain remains in fixative, because in that case, there will be a long-term, dynamic process of covalent crosslinking and loss of crosslinking bonds. As a result, stability is better assessed in specimens that have been removed from fixative. It was shown early in the era of formaldehyde that gelatin fixed with excess formaldehyde forms an insoluble gel, which does not change structure even after being exposed to water for 10 months (Hardy, 1899). One study using radiolabeled formaldehyde found that a portion of formaldehyde-collagen bonds cannot be removed even after up to 19 weeks of washing (Barker et al., 1980). In this study, the percentage of non-removable formaldehyde reached a plateau at 12-20 %, suggesting these formaldehyde molecules had formed stable formaldehyde-collagen bonds. As another example, glutaraldehyde crosslinked heart valves can last for 12-15 years in vivo prior to structural degeneration, which occurs either due to calcification or tearing at suture points (Tam et al., 2017). As a result, there is reason to think that aldehyde-based crosslinking bonds can be stable for long periods of time. However, this is a complex question that undoubtedly depends on the context. For example, the stability of crosslinks depends on the concentration of aldehyde used prior to the removal from fixative (Barker et al., 1980; Lyon et al., 1991). Additionally, the stability of the crosslink depends on the type of chemical bonds formed between formaldehyde and the crosslinked biomolecules (Gavrilov et al., 2015; Kamps et al., 2019). Other factors include the fixative used, the tissue type, and the storage medium. Indirectly entangled biomolecules likely include lipids such as phosphatidylcholines and sphingomyelin that are not directly crosslinked by fixatives but appear to be retained as a part of the membrane protein-lipid complex (Denti et al., 2020). If a biomolecule is indirectly entangled in the chemical gel, then there are a few possible ways that its location could change during long-term storage. Namely, the biomolecule could (a) become chemically modified and covalently bound to the mesh by residual aldehyde, (b) remain confined long-term in the absence of a covalent bond, (c) diffuse to a significantly different location within the mesh, or (d) leak out of the mesh and dissolve in the solvent. We can think of an entangled biomolecule as a guest molecule embedded in a chemical gel matrix (Arends et al., 2015; Kowalczuk et al., 2016; Chen and Muthukumar, 2021). In certain scenarios, the biomolecule may become effectively immobilized due to barriers that hinder its diffusion (Chen and Muthukumar, 2021). The propensity of a guest molecule to disperse is an empirical question dictated by multiple factors, including its radius of gyration relative to the local mesh size, the gel's biochemical composition, non-covalent interactions such as electrostatic bonds, crosslink density, and the temperature. For instance, during aldehyde fixation, triglycerides, cholesterol, and glycogen may remain physically trapped without direct crosslinking (Lyon et al., 1991). Aldehyde storage After the initial step of fixation in the brain preservation procedure, the most straightforward and common method for long-term fluid preservation is to simply leave the brain in the same fixative solution (Feirabend and Ploeger, 1991; Vonsattel et al., 2008; Kövari et al., 2011). Some previous authors have suggested that storage can be effective for the long-term in formaldehyde solutions. For example, two sources note that brains can be stored in formalin for an indefinite period (Fish, 1895; Voogd and Feirabend, 1981). However, even though brains can be stored for long periods in solutions containing formaldehyde, the key question is the quality of the preservation over time, such as the degree to which specific features can still be distinguished on microscopy as expected with a given visualization technique (Koehler et al., 2024). Therefore, understanding the empirical effects of this storage method is essential. Empirical studies of formaldehyde storage We built a database of studies measuring the effects of formaldehyde storage on cellular morphology in nervous system tissue, employing a wide range of methodologies (Supplementary File 7; example results in Figure 3). Observations extracted from these studies were independently graded by two raters for storage artifact severity on a subjective 0-2 scale, with “0” indicating no or minimal artifact, “1” partial, and “2” severe or total. Inter-rater reliability for these grades was excellent, as indicated by an intraclass correlation statistic of 0.958 (F-test p-value = 8.9 * 10-70) (Koo and Li, 2016). Among the 155 observations, 60.8 % reported no or minimal storage artifact, 22.2 % a partial storage artifact, and 17.1 % a severe or total storage artifact (Figure 4). We identified one type of biomolecular artifact – loss of antigenicity (n=47 observations) – and five types of morphological artifacts: decreased silver staining (6.9 %), decreased structural preservation (n=2 observations from (Lai et al., 2018)), areas of empty neuropil (n=2 observations from (van Duijn et al., 2011)), myelin-like whorls (n=2 observations from (Robards and Wilson, 1993)), and nuclear degeneration (n=2 observations from (Cook et al., 2014)). We first discuss the biomolecular alterations and then the morphological alterations observed during storage in formaldehyde solutions.
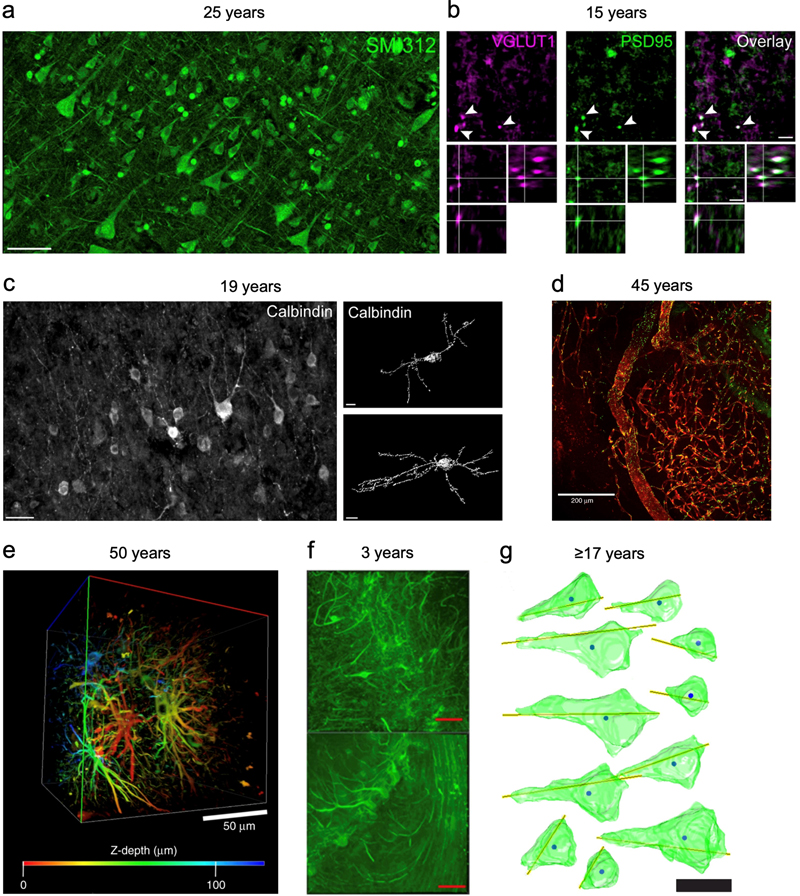
Figure 3. Example images showing morphology preservation from tissue stored long-term in solutions containing formaldehyde.
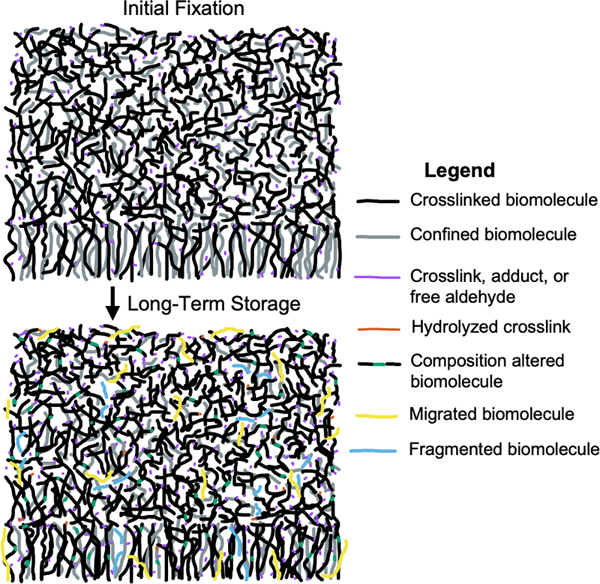
Figure 4. Severity grades for effects of formaldehyde storage on cellular morphology in brain tissue. Effects on proteins The effect of fixative storage on proteins depends strongly on the protein and the techniques used for detection (Thacker et al., 2021). In immunohistochemistry, the duration of fixation needs to be timed precisely for optimal staining. Underfixed tissues have altered tissue morphology and poor antigen retention, while overfixed tissues have poor antigen staining, largely because excessive crosslinks hinder antibody penetration and binding to antigens (Beckstead, 1994). Numerous studies found that there is a loss of staining for certain antigens following storage in formaldehyde for months, years, or decades (Beach et al., 1987; Dwork et al., 1998; Sheaffer et al., 1999; Pikkarainen et al., 2010; Lundström et al., 2019; Wu et al., 2022; Lin et al., 2023). For example, one study found that fixation times need to be precisely controlled when staining formaldehyde-sensitive antigens such as DCX, PSA-NCAM, and NeuN (Flor-García et al., 2020). In this study, fixation times of more than 12 hours were shown to have strong effects on staining properties, and staining could be completely abolished with more than 6 months of fixation time. Antigen retrieval methods can often remove excessive crosslinks and allow for binding of antibodies to proteins despite long durations of fixation (Evers and Uylings, 1997; Liu et al., 2010). For example, one study found that tissue clearing using CLARITY led to substantially increased immunostaining for CAMKIIA in a brain that had been fixed in formalin for 18 years (Woelfle et al., 2022). The authors speculated that this was because the clearing led to mild disruption of highly crosslinked formaldehyde-protein networks, thus making the epitopes more accessible. However, there are also limits to current antigen retrieval methods. One study of brains stored in formalin for up to 14 years found that while antigen retrieval helped to improve the visualization of some antigens, other antigens still had diminished or absent staining as a result of storage (Pikkarainen et al., 2010). Other sources have used unbiased profiling methods to analyze numerous proteins at once. One study used proteomics following antigen retrieval techniques, finding that mouse brain tissue fixed for 3 years did not yield more proteins than human brain tissue fixed for 7 years (Rahimi et al., 2006). Indeed, they found that human tissue fixed for 7 years detected a similar number of proteins as was detected in fresh frozen human brain tissue (Rahimi et al., 2006). Another study developed a method of mass spectrometry on fixed tissue following brain clearing (Bhatia et al., 2022). They compared tissue that had been formalin fixed for more than 5 years to PFA fixed control samples, with more than 5000 proteins identified, and Pearson correlations between the proteomes from these two conditions ranging from 0.91 to 0.96. This result suggests that overall protein content is largely maintained during storage in formaldehyde, for at least 5 years. Notably, other studies have reported biochemical changes in proteins as a result of formaldehyde fixation; for example, hydrolysis of certain proteins is thought to occur (Matsuda et al., 1998; Hackett et al., 2011). However, the extent of these chemical changes remains unknown. Taken together, extant data shows that most proteins remain present in fixative-stored brain tissue over a timescale of years, albeit potentially difficult to access with antibodies and with the potential for chemical alterations. Effects on lipids One early study investigated the effect of formalin storage on the ability to extract and visualize lipids with chromatography from several types of animal tissues, including brain and spinal cord (Heslinga and Deierkauf, 1961). They found that phosphatidylethanolamine was no longer visualized after 93 hours of fixation. In samples fixed for more than one year, there were further changes, including a reduction of lecithin. On the other hand, the levels of other lipids, such as cholesterol and sphingomyelin, were not substantially affected. The authors proposed that the observed changes might result from modifications in the chromatographic and staining characteristics of the lipids, or possibly due to changes in lipid extractability after they had more time to interact with tissue proteins. A more recent study using mass spectrometry found that there was a loss of phosphatidylethanolamine and phosphatidylserine detection in fixed brain tissue, while sphingolipids remained intact after years of storage in formalin (Gaudin et al., 2014). On this basis, they suggested that phospholipids are largely degraded by hydrolysis, oxidation, and covalent modifications. However, a later study argued that their result was not due to the chemical degradation of the phospholipidome, but rather that the aminophosholipids are rendered inaccessible to their method of mass spectrometry, which is a rapid effect that occurs after only overnight fixation (Carter et al., 2016). Consistent with the notion that many lipids are not chemically degraded during fluid preservation, a recent study used mass spectrometry imaging on fresh frozen and formalin-fixed brain samples to detect gangliosides, which are a type of sphingolipid often found on the surface of neurons (Harris et al., 2020). They found that although immersion in formalin initially decreases lipid signal compared to fresh frozen tissue, there was a similar signal enhancement in rat brains fixed for 15 minutes and human brains fixed for up to 15 years, suggesting that long-term storage in formalin does not significantly affect ganglioside content. Notably, following long-term storage in fixative for many decades, brain tissue has been reported to be “virtually non-clearable” (Lai et al., 2016). This suggests that long-term chemical modifications may cause a subset of lipids that are at first indirectly confined to become more strongly bound to the crosslinked biomolecular mesh over time. In addition, it has been speculated that the degree of chemical alterations to lipid species, such as hydrolysis and oxidation, likely correlates with the storage duration of specimens in formalin (Dannhorn et al., 2022). Overall, there may be significant chemical alteration of tissue lipids when brain tissue is stored in fixative for the long term, but it seems that a substantial subset of lipids is retained. Effects on nucleic acids Formaldehyde is well-known to cause chemical changes in nucleic acids, such as cytosine deamination and depurination, which become more common over time (Raxworthy and Smith, 2021). The ability to sequence nucleic acids is rapidly lost during the initial days of the fixation process, because sequencing relies on dissociating the molecules, and crosslinking prevents this (Guo et al., 2023; Vitošević et al., 2023). However, nucleic acid profiling is still possible after many decades or even more than a century. For example, one study sequenced 1918 pandemic influenza RNA from samples fixed in formalin for close to a century, after using heat treatment to partially reverse the formaldehyde crosslinks (Patrono et al., 2022). In addition, one study found that the distribution of DNA sizes extracted from brains stored in formalin was not altered when comparing storage times of 3 years up to 46 years, suggesting that long-term storage does not lead to a linear decrease in molecular preservation quality (Savioz et al., 1997). As with proteins and lipids, extraction methods play a critical role in determining the success of profiling nucleic acids in long-term formalin-fixed tissue (Herbin et al., 2021). For example, one study found that there was a decreased in situ hybridization (ISH) signal for DNA following fixation in 7 % neutral buffered formalin for 79 weeks (Mostegl et al., 2011). However, through an increase in the concentration they used of proteinase K – which digests crosslinks and renders the DNA more accessible – prior to performing the ISH assay, the signal returned to the same level as day one of fixation time. Another study found that miRNAs can be profiled in brain tissue that has been stored in formaldehyde for more than 20 years (Herai et al., 2014). Mechanistically, acidic conditions in the tissue, attributed to formaldehyde degradation into formic acid and the degradation of fats into fatty acids, are thought to accelerate DNA hydrolysis, leading to the loss of nucleobases from the DNA molecule (Kösel and Graeber, 1994; Herbin et al., 2021). DNA sequencing in archival tissue is a highly active area of research. Emerging methods have had success in extracting longer DNA molecules from long-term fixed tissue, suggesting that much of the DNA content remains present in cells and tissues even after long-term storage in fixative, and that this can be accessed with the proper techniques (Savioz et al., 1997; Fang et al., 2002; Hykin et al., 2015; Hassani and Khan, 2015; Hahn et al., 2022). Effects on small molecules Regarding small molecules, they are liable to leech out of the fluid preserved brain tissue over time. For example, levels of the cocaine metabolite benzoylecgonine were found at high levels in the formalin solution after 30 d of storage of several tissues in formalin (Hilal et al., 2009). In this study, the leaching was the smallest in brain tissue compared to the other tissues tested, but it is likely still present. Bioelements also can shift substantially during fluid preservation. Tissues fixed with buffered formalin and profiled with mass spectrometry imaging show a shift towards sodium adducts compared to potassium adducts in fresh tissues, likely due to the sodium content of buffered formalin solutions (Carter et al., 2011). One study investigated the levels of 19 elements in brains stored in formalin for approximately 20 years (Gellein et al., 2008). They found that there was a substantial leaching out of some elements, such as As, Cd, and Mg. However, the concentration of most of the bioelements was still much higher in the brain tissue than in the formalin solution they were stored in. Bioelements that are known to be strongly bound to the sulfhydryl groups found in proteins, such as Ag, Hg, and Ni, leached out from the tissue less than others. Summary of storage effects on biomolecules In summary, some chemical species that are not directly attached to the formaldehyde-induced crosslinking meshwork, small molecules and bioelements especially, appear to slowly migrate out of the tissue over time (Figure 5). But the preponderance of data suggest that most biomolecules become increasingly trapped in the formaldehyde meshwork over time. This inhibits our ability to visualize the molecules, leading to the phenomenon of overfixation. There are limits in the extent of clearing and antigen retrieval that may be possible with contemporary technology, but this may improve in the future, further “unlocking” these biomolecules within archival tissue stored in formaldehyde solutions (Thacker et al., 2021; Hahn et al., 2022). Additionally, the biomolecules may undergo chemical reactions such as oxidation and hydrolysis that can alter their chemical composition.
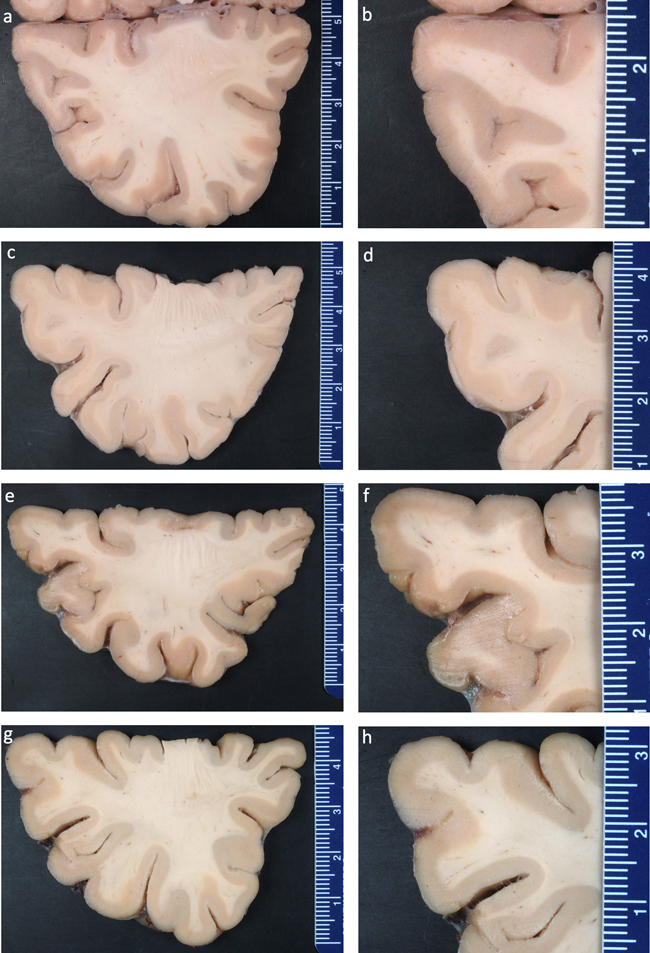
Figure 5. Conceptual diagram of the dynamic changes in biomolecules during long-term storage in fixative. Effects of formaldehyde storage on tissue morphology The most severe storage artifact we identified was in van Duijn et al. 2011, who reported numerous areas of white discoloration on gross examination and corresponding hypointensities on MRI (van Duijn et al., 2011). No such areas were identified in brains stored in formalin for up to 1 year, but all brains stored over 6 years showed them, with increasing frequency with longer storage. These areas are on the order of hundreds of micrometers in size. On light microscopy, they contain granular, basophilic neuropil changes with some tissue rarefaction, and decreased Kluver's staining, thereby indicating a lower density of myelin. The authors noted that cells and vessels are unaltered in these areas. On electron microscopy, these areas contain spaces with absent or minimal neuropil, and varying amounts of lamellar structures, interpreted as membrane remnants and/or degenerated myelin sheaths. Notably, the brains in this study were stored in sealed plastic bags with a small excess of 10 % formalin, as opposed to the more common method of storage immersed in fluid in a glass or plastic container. The storage artifacts identified in van Duijn et al. 2011 were found throughout the cortex, and should be detectable by many other studies, including as white discolorations on gross examination, hypointensities on MRI, areas with basophilic neuropil on light microscopy, and localized empty spaces on electron microscopy. However, other studies did not report these artifacts, including other electron microscopy studies (Dykstra, 2010; Liu and Schumann, 2014; Tsutsumi, 2018), and studies that could account for sampling bias by using neuroimaging (Herbin et al., 2021; Wiggermann et al., 2023). As an additional test, we grossly examined several brain samples stored via fluid preservation in the Neuropathology Brain Bank at Mount Sinai for between 3-5 years and we did not identify any white discolorations (Figure 6). Taken together, we suggest that the neuropil decomposition this study found during storage may be associated with the plastic bag storage method that likely leads to more air exposure, while also pointing out that this is a critical area for further research.
Figure 6. Gross examination of postmortem human brain tissue reveals an absence of macroscopic discolorations in the cortex. One study found that storing rat brain tissue in 2 % paraformaldehyde and 2.5 % glutaraldehyde buffered by sodium cacodylate for 1 month led to more frequent intracellular and extracellular “myelin-like” whorls on electron microscopy than were seen after 3 hours of fixation (Robards and Wilson, 1993). By 12 months, the whorls appeared less membranous and were associated with areas of lucency, suggestive of lipid leaching. The authors suggest that this artifact may be due to incomplete fixation of phospholipids, causing them to dissolve, coalesce, and then take on this myelin-like appearance after reacting with osmium tetroxide. They note that the preservation of cellular organelles was stable with long-term storage and that the length of fixation is thought to be less critical for morphology than other variables like the fixative used, the buffer, the pH, and the osmolarity. Another study found that similar myelin-like whorls were more common inside and outside of cells in skin tissue on electron microscopy after storage for 4 weeks in 2.5 % glutaraldehyde (Lindberg, 1984). Myelin-like figures have also been associated with mitochondrial degeneration (Le Beux et al., 1969). It has been speculated that they result from the reaction of extracted phospholipids with osmium tetroxide (Ericsson et al., 1965). Unless they become severe, the myelin-like whorl artifacts seem to be small enough that they would be unlikely to substantially affect the mapping of neural circuitry. In addition to these morphological artifacts, there are several others to consider. One study notes that nuclei degrade in brain tissue stored in formalin and that this progressively worsens from 15 to 80 years of storage (Cook et al., 2014). The authors suggest that for the samples fixed for a longer period, different formulations of formalin may have been used, such as a non-buffered version. In contrast, other studies did not describe a loss of nuclei even after many decades of storage in formaldehyde (Sheaffer et al., 1999; Herbin et al., 2021). Several studies note that silver staining can be altered following extended formalin fixation (Wilcock et al., 1990; Heinsen et al., 2000; Rosoklija et al., 2003). However, this appears to be dependent on the method of silver staining. For example, one study notes that the rapid Golgi method gave inadequate results, while the Golgi–Kopsch method yielded excellent quality staining comparable to tissue fixed for a few years, even on tissue stored in formalin for up to 55 years (Rosoklija et al., 2003). One study that used cresyl violet staining to visualize neuronal morphology reported that “structural preservation” was not as good in tissue fixed in formalin for many years, without providing more detail on what aspect of the structure was altered (Lai et al., 2018). As in immunostaining studies, overfixation might help to account for decreased morphologic staining intensity after prolonged fixation. Because morphological stains non-specifically bind to many types of biomolecules, if there is a global decrease in biomolecular binding capacity due to excessive crosslinks, then this will lead to a global decrease in the signal to noise ratio of morphological staining. Some biomolecules may also be extracted from cells over time, making the cytoplasm of cells appear more pale (Lindberg, 1984; Robards and Wilson, 1993). Additionally, because long-term fixation can render tissues more acidic, the staining quality of anionic dyes may be especially decreased (Puchtler and Meloan, 1985). In addition to histologic effects, long-term storage in fixative can also have macroscopic and biomechanical effects. Upon initial fixation, fixatives often cause cell shrinkage, which is accentuated for stronger fixatives such as glutaraldehyde (Nei, 1981). One source finds that after the initial fixation, the volume of canine brain structures was not altered due to additional storage in formalin for up to 12 months (Del Signore et al., 2022). On the other hand, other reports have suggested that specimens do have secondary shrinkage if they are stored in fixative fluids for long periods (Bahr et al., 1957; Heinsen et al., 2000). This discrepancy may be due to the type of brain tissue, the fluid properties, and the duration of storage. Macroscopic shrinkage may correspond to widespread microscopic strengthening of chemical gels in the tissue over time. Strengthening of microscopic interactions, however, is associated with changes in the material properties of the brain, insofar as it becomes more stiff, brittle, and difficult to section (Dannhorn et al., 2022). Prolonged storage in fixative can also promote the accumulation of formalin pigment (Sacchini et al., 2022). Formaldehyde formulation The concentration and formulation of formaldehyde may play an important role in long-term storage. Several studies have used a lower concentration of formaldehyde for storage, such as 0.1 % paraformaldehyde (Lyck et al., 2008), 1 % formalin (Papageorgopoulou et al., 2010; Wiggermann et al., 2023), 2 % paraformaldehyde (Wu et al., 2002), or 2 % formalin (Monteiro, 2008). Using a lower concentration of formaldehyde during storage is appealing because it may help to prevent the negative effects of formaldehyde while still preventing microbial growth and any theoretical loss of morphologic cohesion. In terms of additives, there is a concern that storage in commercial formalin solutions, which contain a small amount of methanol in addition to formaldehyde, may lead to lipid damage, because methanol is a solvent for lipids (Öztürk and Koç, 2022). However, one study found that even after 18 years of storage in methanol-stabilized formaldehyde, staining with a lipophilic dye was still possible, and that there was no difference in the distance of nerve tracing possible as a result of storage (Öztürk and Koç, 2022). Storage in other aldehydes In addition to formaldehyde, other aldehyde fixatives can also be used for long-term storage. Although this is less well studied, these storage solutions are expected to have many of the same effects as formaldehyde. Tissue stored in other aldehyde fixatives will undergo overfixation. Other aldehyde chemicals also have the potential to convert to acid forms. For example, concentrated glutaraldehyde oxidizes to form an organic acid during storage, which has been found to decrease the pH of the solution over time (Boucher, 1978; Ranly, 1984). When glutaraldehyde is used in the fixation of nerves, studies have reported switching the solution to formaldehyde or phosphate buffer for long-term storage (Magoon and Robb, 1981; Morgello et al., 2001). One source notes that nervous system tissue “deteriorates rapidly” in glutaraldehyde; however, they do not specify the outcome measure they used to make this evaluation (Voogd and Feirabend, 1981). Because glutaraldehyde is a stronger fixative than formaldehyde, problems of overfixation and inaccessibility of biomolecules are expected to occur more quickly during storage in glutaraldehyde. However, one source reports that storage in glutaraldehyde is compatible with good long-term structural preservation, with no evidence of ultrastructural alterations to kidney tissue stored in 4 % formaldehyde and 1 % glutaraldehyde at 4 °C for up to 23 years (Dykstra, 2010). One publication reported using glyoxal for long-term storage, but did not report the outcomes of tissue stored in this manner (Thomas et al., 2022). Summary The most common storage artifact identified in the studies reviewed is a loss of antigenicity for certain proteins over time, likely due to the accumulated crosslinking making antigens inaccessible. While some antigens become undetectable after days, months, or years of storage, other proteins can still be visualized even after decades in fixative. Other biomolecules like nucleic acids and a subset of lipids also frequently become more heavily crosslinked and confined over time, making them more difficult to profile, but usually not impossible to profile with specialized techniques. Besides effects on biomolecules, morphological artifacts were less frequently reported. One study found localized empty spaces in neuropil and after more than six years of storage, but this finding was not reported by other studies with similar or longer storage times, raising the question of whether the severity of this artifact was associated with their storage method in plastic bags (van Duijn et al., 2011). One study noted the accumulation of “myelin-like” whorls during storage that was associated with incomplete fixation of lipids and lipid leaching (Robards and Wilson, 1993). Another study reported nuclear degradation after decades of storage, but others did not note this effect (Cook et al., 2014). More commonly, nonspecific decreased staining intensity was noted in some studies, potentially also related to chemical alterations of biomolecules. Overall, while there have been some storage artifacts reported following-long term storage, the extent to which aspects of the underlying neural circuitry information is irreversibly lost, or simply more difficult to measure, as well as how this varies between different fixative storage methods, are open questions worthy of further investigation. Buffer storage Buffer solutions such as sodium phosphate buffers, phosphate-buffered saline (PBS), and cacodylate buffer are commonly used for maintaining a stable pH during the preservation of fixed brains (Supplementary File 8). PBS is designed to be an isotonic, non-toxic solution that mimics the extracellular fluid, and is among the most common buffers used in the field of brain banking. Cacodylate buffer is a heavy metal buffer commonly used in electron microscopy that avoids the risk of calcium phosphate precipitates and microorganism contamination that can occur with the use of phosphate buffers (Ivanchenko et al., 2021). A common method for long-term fluid preservation of fixed brain tissue is to transfer brains to a buffer solution, without any fixative. As discussed above, the pH buffering capacity of these solutions can diminish over time, rendering long-term storage in buffers essentially equivalent to storage in water. Two other factors that are important to consider is the degree of residual aldehyde fixative remaining in the solution and the prevention of microbial contamination. Degree of residual fixative One option to decrease the amount of formaldehyde in the solution is to attempt to wash it out. However, even washing formaldehyde fixed tissue for weeks has been found to be insufficient to remove all formaldehyde crosslinking bonds (Barker et al., 1980). Washing tissue is an even a greater problem in the study of intact whole brains because of their size. For example, washing human brains with buffer for 48 hours has been found to be clearly insufficient for buffer penetration to interior brain regions, leading to a contrast boundary artifact (Miller et al., 2011). As a result, despite the imaging artifacts associated with formaldehyde, washing fixed whole human brains is often not performed prior to neuroimaging because removing formaldehyde would take a prohibitive amount of time (Tendler et al., 2021). One source suggests that washing tissues in water for an extended number of years does eventually dislodge more formaldehyde (Puchtler and Meloan, 1985). Because washing fixed tissue for extensive periods of time can still yield formaldehyde in the effluent, it is tricky to distinguish the bound formaldehyde from that which can be removed (French and Edsall, 1945). However, there appears to be thresholds beyond which additional washing does not yield a substantial amount of additional fixative (French and Edsall, 1945). Effectively, storage in buffer constitutes storage in a much lower concentration of fixative solution, with the concentration depending on the amount of time that the tissue was washed prior to long-term storage. Preventing microbial growth One of the major potential downsides of removing the brain from fixative storage is the potential for microbial growth. One source reports that this is a common occurrence in buffer solutions, as proxied by bacterial or fungal growth in the absence of a host immune reaction (Dykstra, 2010). As a result, it is commonly thought that storage in buffer requires adding an additional chemical agent to prevent microbial growth and resulting tissue damage. The most common agent for this is sodium azide (NaN3), an antimicrobial preservative that acts to inhibit mitochondrial respiration, largely by binding to cytochrome c oxidase (Ishii et al., 2014). For example, one source reports that if fixed brain tissue is stored in PBS at 4 °C for longer than a week, 0.02 % of sodium azide should be added to the PBS solution to prevent bacterial growth (de Prisco et al., 2022). Another study reports that the addition of sodium azide can help with maintenance of histochemical staining in brain sections (Morán and Gómez-Ramos, 1992). Thymol is another antimicrobial agent that can be used, which acts to disturb the integrity of bacterial and fungal membranes (Hammer et al., 2012). The question of how long sodium azide retains its antimicrobial activity during storage is not well established in the literature. Notably, sodium azide is used at a range of concentrations in preservation solutions; for example, it has been used at 0.1 % (Beach et al., 2015) or 0.01 % (Micheva et al., 2023). However, concentrations lower than 0.01 % may not allow for effective antimicrobial activity (Snyder and Lichstein, 1940). At relatively higher concentrations, there is more leeway against the potential for sodium azide to break down chemically over time and no longer protect against microbial growth. Effects on cellular morphology One of the major advantages of buffer storage is in mitigating or preventing overfixation, which damages antigenicity and makes sections difficult to process. One brain bank reports that they store fixed brain blocks in 0.1 mol/L phosphate buffer with 0.1 % sodium azide at room temperature, with excellent results for immunohistochemistry studies after years of storage (Beach et al., 2015). One study stored brain tissue for around 1.5 to 15 months in 0.1 M cacodylate buffer pH 7.4 or 0.1 M phosphate buffer with sodium azide at 4 C prior to embedding for electron microscopy (Sele, 2020). They reported that there was no difference in the ultrastructural preservation quality when comparing tissue that was stored for this length of time or not, suggesting that there were no adverse effects of the storage on tissue quality. Storage in phosphate or cacodylate buffer also has been recommended for storage of nerve tissue fixed with glutaraldehyde (Morgello et al., 2001). Finally, phosphate buffer has been used as a control storage medium when studying brain clearing methods (Furuta et al., 2022). Overall, because it is not as common of a method, not as much long-term preservation data is available, but the available data suggests that buffer storage is not associated with damage to neural structures over the time scale of years. Effects on brain volumes A decreased effect on brain volumes is thought to be an advantage of fluid preservation in phosphate buffer compared to storage in formaldehyde, which has been associated in some studies with long-term shrinkage due to cross-link formation and the partial leaching of soluble solids (Hackett et al., 2011; de Guzman et al., 2016). This might be useful for studies attempting to measure the volume of a particular brain region. However, the effect of buffers on brain volume sizes depends on the osmotic concentration. One study found that five months of storage of fixed brains in a standard concentration of PBS led to a slight decrease in volume in several brain regions (de Guzman et al., 2016). On the other hand, three weeks of storage in distilled water led to increases in volumes across the brain, while three weeks of storage in 10x PBS led to a dramatic shrinkage in volumes across the brain (de Guzman et al., 2016). Summary Buffer solutions are commonly used for long-term storage of fixed brain tissue. A major advantage is preventing overfixation and associated loss of antigenicity. However, washing fixative from large brains is challenging, so residual aldehyde may remain. Microbial growth also needs to be prevented, which is often performed using sodium azide. There is theoretical reason to expect that buffer storage would prevent overfixation and morphological alterations associated with formaldehyde storage, but long-term data is limited. The long-term breakdown of the stabilizing chemical gel without additional crosslinking agents is also a theoretical concern. Taken together, buffer storage helps address some limitations of aldehyde storage, but open questions remain about very long-term preservation quality. Alcohol storage Alcohols, especially ethanol, have long been recognized as a useful medium for fluid preservation (Supplementary File 9). Ethanol is still commonly used today in museum collections, where a low concentration of formaldehyde is often used for initial fixation, followed by long-term preservation in ethanol (Straube et al., 2021). The use of other alcohols like methanol and isopropanol has also been documented, but ethanol tends to be the most used due to its relatively lower toxicity. A notable concern is the flammability of alcohol, which requires careful storage procedures to mitigate safety risks. Mechanisms The main mechanism by which alcohols act as preservatives is to dehydrate tissues, eliminating water from cells and consequently leading to protein denaturation and prevention of enzymatic degradation. In this context, it is sometimes called a coagulative fixative. It also possesses antimicrobial properties, which helps to prevent contamination of stored specimens. The elimination of water can lead to severe shrinkage of the tissue, affecting morphology. Many investigators use an ethanol concentration of around 70-80 % for the long-term preservation of fixed brains, balancing preservation efficacy with less damage to tissue morphology. However, the optimal concentration of ethanol depends heavily on the context and desired use, with more morphological damage but less biomolecular damage occurring at higher concentrations (Marquina et al., 2021). Effects on biomolecules In contemporary use, alcohols are sometimes favored over aldehydes as a preservative due to their less damaging effects on certain classes of biomolecules, mainly DNA and proteins. For example, the yield of DNA from brain specimens was altered after one month of storage in formaldehyde, but not after the same duration of storage in 75 % ethanol (Zamenhof et al., 1972). More recent studies have confirmed that spirit-preserved specimens have higher quality DNA when stored in ethanol as compared with formaldehyde, although contemporary retrieval techniques following formaldehyde storage have improved substantially (Straube et al., 2021; Hahn et al., 2022). Regarding proteins, one study of preserved fish found that specimens initially fixed in formaldehyde and then transferred to isopropyl alcohol or ethanol for storage for many years (up to 70) had high-quality histology (Kwan et al., 2022). They found that specimens preserved in formaldehyde alone for long periods of time had higher autofluorescence and challenges with achieving high histologic quality. The likely reason that alcohols are less damaging to DNA and proteins is likely that they avoid the overfixation that occurs during storage in aldehydes (Zamenhof et al., 1972). Ethanol has also been used as a storage medium after 24 hours of formaldehyde fixation to prevent overfixation during transport (Stumm et al., 2012). Although alcohol preservation avoids overfixation, it comes with the disadvantage of extracting certain classes of biomolecules from the tissue, especially lipids. Ethanol can dissolve lipids out of the specimen, which can cause them to accumulate as fatty acids at the top of the storage container (Hartman, 2019). This can make the preservative solution cloudy (Stoddart, 1989). The increased acidity due to the accumulation of fatty acids is also thought to contribute to specimen damage (Straube et al., 2021). Due to its denaturing properties, ethanol storage can also damage the conformation and fluorescence of proteins, such as green fluorescent protein (Becker et al., 2012). Effects on morphology Evaluating the literature on fluid preservation with alcohols, particularly ethanol, is challenging due to its historical use as an initial fixative rather than solely as a long-term preservative following formaldehyde fixation. As fixatives, alcohols are generally less effective than crosslinking aldehydes at initial fixation, often resulting in lower quality morphological preservation. For example, one study that perfused 70 % ethanol to preserve a human brain found that it did not harden and displayed a red coloration even two months after fixation, suggestive of inadequate preservation (Grinberg et al., 2008). Another study found that brains preserved in formaldehyde for decades have much better histology than brains preserved in ethanol (Herbin et al., 2021). In this study, tissue preserved in ethanol alone was found to have numerous holes when visualized by light microscopy. Investigations using neuroimaging provide additional context. For instance, spiders stored in ethanol for 70-90 years showed no macroscopic changes in neural architecture, with the exception of two specimens that appeared to have suffered from desiccation (Rivera-Quiroz and Miller, 2021). This is consistent with a study on a human brain preserved in a German collection for 150 years, which also showed satisfactory preservation at the macroscopic level (Schweizer et al., 2014). However, a more in-depth MRI study of this collection identified the presence of sub-millimeter tubular cavities in the white matter of human brains preserved in ethanol for long periods of time (Helms et al., 2014). These tubular cavities were not seen in a bovine brain preserved in ethanol for four months, suggesting that they are an artifact of long-term storage (Helms et al., 2014). These findings led the authors to suggest that ethanol preservation is associated with rapid water and cholesterol loss, followed by longer-term macroscopic deterioration that could be due to a slow loss of lipids. However, ethanol might serve much more effectively as a long-term preservative following fixation. As discussed above, studies in non-brain tissue have found that immunohistochemistry is well-preserved following storage in ethanol as a secondary agent (Stumm et al., 2012; Kwan et al., 2022). One early source suggested that ethanol could serve as an appropriate storage medium for museum specimens following formalin fixation (Fish, 1895). Even prior to the use of formaldehyde in pathology, one source noted that if brain tissue was not adequately hardened in Muller's fluid (a dichromate solution acting as a fixative), then long-term preservation in strong alcohol of greater than 80 % could lead to significant damage (Van Gieson, 1889). Specifically, the alcohol would dissolve out the fixative salts and alter the myelin over a period of 1-2 years. This process would result in the formation of cholesterol crystals in the alcohol, and microscopic cavities or vacuoles in the specimens, which did not stain well. Van Gieson noted that even if the tissue were adequately hardened, there would still be problems with histology after storage in alcohol, such as neuroglia not being visualized well, compared to tissue stored in water that had a low concentration of Muller's fluid. Another source notes that storage of fixed tissues in alcohol over just one weekend in can lead to extensive vacuolization of white matter, although this likely depends on the concentration used (Garman, 2011). A different source reports that tumor tissue can be stored in 70 % ethanol for up to 4 weeks with no significant changes of histology and biomolecular staining patterns (Stumm et al., 2012). Finally, one source notes that fixed sperm cells tend to have membrane damage when they are stored in ethanol without secondary fixation in osmium, consistent with the known role of ethanol in lipid extraction (Fahey and Healy, 2003). Summary Alcohol storage is a cost-effective, practical, and time-tested method for preserving fixed brains. Storage in alcohol has several advantages, including antimicrobial effects and avoiding overfixation. However, it can cause several problems, including tissue shrinkage leading to altered morphology, biomolecular extraction, cavity formation, and vacuole formation. There is sometimes thought to be a trade-off, wherein storage in ethanol has worse morphological preservation, but better biomolecular preservation, compared to storage in formaldehyde. However, that trade-off is likely a practical one based on our currently available biomolecular profiling technology, rather than fundamentally driven by the underlying biochemistry. Overall, following the development of buffer solutions and antimicrobial agents such as sodium azide that can prevent overfixation without causing biomolecular extraction, alcohols seem relatively less useful than they were in the past. They are not commonly used in brain banking and would not be ideal for use in brain mapping studies. However, the extant literature on this topic is not extensive, making it difficult to draw firm conclusions. Storage after tissue clearing Brain tissue clearing is primarily used for imaging intact brain structures in detail by removing parts of the tissue that interfere with 3D visualization, such as lipids and calcium, thus rendering it more transparent (Ueda et al., 2020). Tissue clearing processing procedures are often quite harsh, involving powerful solvents and often high temperatures. For example, the SWITCH protocol uses temperatures of 80 °C for up to 4 days in tissue fixed with glutaraldehyde, equivalent to slightly more than 2 years of storage at 4 °C, assuming a Q10 value of 2 used in shelf life studies (Murray et al., 2015). Although not primarily developed for long-term storage, several types of brain clearing techniques have also been reported to provide high-qualitylong-term storage of brain tissue, so they merit consideration as fluid preservation methods (Supplementary File 10). Empirical effects of brain clearing There are three classes of brain clearing approaches, each with different effects on preservation quality: hydrophobic, hydrophilic, and hydrogel methods (Ueda et al., 2020). Hydrophobic clearing methods use organic solvents to clear the tissue, which hardens them and allows for long-term preservation, re-analysis via immunolabeling, and stabilization of endogenous fluorescent signals. Notably, hydrophobic clearing methods sometimes use ethanol as a dehydrating agent, but also use additional steps for delipidation and refractive index matching, thus differentiating these techniques from fluid preservation using ethanol on its own. One example of a hydrophobic brain clearing approach is SHANEL, which has been reported to allow for long-term storage and future histologic studies of the same tissue (Zhao et al., 2020). As another example, sDISCO has been found to retain structural details and protein fluorescence after storage for 22 months (Hahn et al., 2019). Methyl salicylate is another hydrophobic reagent that is the active ingredient of wintergreen oil and was a component of the first clearing method used by Spalteholz in 1911 (Liu et al., 2016). One study found that storage of fixed human choroid plexus in methyl salicylate for up to 25 years allowed for high-quality immunohistochemical staining of several antigens (Sufieva et al., 2023). Another study found that methyl salicylate combined with permanent mounting allowed storage of insect brains for well over a year at room temperature with intact immunohistochemistry (Bekkouche et al., 2020). In hydrophilic clearing methods, reagents can be used for decolorization, delipidation, and refractive index matching, but water is not removed from the specimen (Ueda et al., 2020). These techniques usually have reagents that form hydrogen bonds with proteins, which can aid in the preservation of protein conformation and associated fluorescence (Ueda et al., 2020). In one study of a modified version of the hydrophilic brain clearing method CUBIC, the authors reported that the samples were washed and stored in PBS with 0.01 % sodium azide in case re-imaging was needed in the future (Pinheiro et al., 2021). Finally, in hydrogel brain clearing methods, chemicals are introduced to strengthen tissue gels, either via introducing molecules to create a synthetic gel, or via epoxy crosslinkers to fortify existing tissue gels (Ueda et al., 2020). For example, in the CLARITY method, acrylamide is used to create a hydrogel-tissue hybrid that aids in retaining proteins and maintaining structural integrity during subsequent processing steps such as delipidation (Malkovskiy et al., 2022). The extent to which the extra stabilization provided by hydrogel methods might improve structural maintenance in long-term fluid preservation of brains is an open question, warranting additional research. Comparison to buffer storage As discussed above, protocols for brain clearing sometimes recommend transferring the specimen back to PBS for long-term storage after clearing (Ke and Imai, 2014). It is interesting to compare this to the buffer storage only approach. Clearing followed by storage in PBS allows for the removal of potentially damaging biomolecules, without the clearing reagents themselves potentially damaging brain tissue over time. For example, lipids are known to be a major source of oxidants during storage of tissue, so removing them could decrease the accumulation of oxidative species during fluid preservation (Huang and Ahn, 2019). Removing lipids also prevents their long-term breakdown into fatty acids and associated decrease in specimen pH. One source reports that the storage of cleared brain tissue in PBS with 0.1 % Triton-X and 0.01 % sodium azide can be performed “indefinitely” (Liu et al., 2016). From the perspective of brain mapping, obvious downsides of delipidation prior to storage in PBS are that the lipids cannot be profiled and the ultrastructure will be dramatically altered (Malkovskiy et al., 2022). Summary Brain tissue clearing techniques like CLARITY and sDISCO have shown promise for long-term storage while enabling intact imaging. Clearing removes light-scattering elements like lipids, which could otherwise damage tissue over time. This is often followed by storage in buffer, avoiding unnecessary further extraction by clearing reagents. Compared to many storage methods relying on embedding in polymers, clearing approaches are more practical, while still achieving many of the same benefits (Sufieva et al., 2023). Though delipidation obviously causes information loss, clearing may better maintain protein structure when compared with other techniques. Clearing techniques also differ in the extent to which they can be practically used in large volumes such as an intact human brain (Zhao et al., 2020). Overall, clearing techniques are an emerging approach that may enable high-quality preservation in addition to allowing intact imaging, but with limited long-term data. Cryoprotectant storage Cryoprotectants, such as glycerol, ethylene glycol, and sucrose, can improve the preservation of biospecimens even above freezing temperatures (Supplementary File 11). While cryoprotectants are best known for protecting against ice damage during subzero temperature storage, we focus here on their biostabilization properties above 0 °C. Indeed, cryoprotectants often act as osmolytes as well, which are small organic molecules that tend to accumulate in cells to help them survive under various stress conditions, such as temperature changes, pressure, and osmotic stress (Rumjanek, 2018). By substituting for the water molecules that typically bind to biomolecules, cryoprotectants tend to increase intracellular viscosity, reduce molecular motion, and slow down reactions, preserving biomolecular integrity without causing significant extraction. They also help preserve hydrophobic interactions and the native structural properties of biomolecules, like protein fluorescence (Nürnberg et al., 2020). Reactions that depend on water, such as hydrolysis, will not occur in pure cryoprotectant solutions, because water is not present. However, chemical reactions not relying on water will still occur, potentially contributing to tissue degradation over time. For example, one study found that when dissolved in a 99.8 % glycerol solution, the enzyme lysozyme still retained one-seventh of its catalytic activity compared to water (Rariy and Klibanov, 1997). Finally, certain cryoprotectants such as sugar alcohols have been found to play a role in stabilizing gel structures for the long-term, consistent with their role in protein stabilization (Baydin et al., 2022). Glycerol Glycerol is a nontoxic cryoprotectant that was used as an early preservative for biological specimens (Van Dam, 2020). Its antiseptic and humectant properties can help slow decomposition. Glycerol began to be used in the late 1800s as an alternative or complement to ethanol for storage in museum collections. Carlo Giancomini used glycerol for storing zinc chloride-fixed and alcohol-dehydrated brains, with good reported efficacy in both dry and fluid preservation (Thomson, 1880). Glycerol also gained use for stabilizing formaldehyde-fixed tissues, with some human organs reportedly stored in this manner with excellent macroscopic condition after more than 100 years (MacLeod et al., 2018). The effects of glycerol on tissue microstructure and morphology are quite variable between studies. It depends on multiple factors like the tissue type, glycerol concentration, how rapidly the concentration is increased, and the degree of hydration (Armitage, 1986; Nowacka et al., 2012). However, many studies have found that gradual increases of glycerol do not cause major structural changes like shrinkage. For example, cell membranes are thought to be the structures most vulnerable to damage during dehydration (Westh, 2003). Glycerol helps preserve cell membranes during dehydration through a mechanism that appears to involve interacting with polar lipid headgroups (Nowacka et al., 2012; Pocivavsek et al., 2011). Consistent with the idea that glycerol is able to maintain cellular structure, one study found that glutaraldehyde and osmium fixed sural nerve biopsies had intact ultrastructure and collagen content after storage in 100 % glycerol at 4 °C for up to 9 months (Myers et al., 1977). The authors stated that tissues could be "stored indefinitely" in this manner. Kaiserling III is a glycerol-containing fluid preservation solution introduced in the late 18th century, with the primary goal of maintaining the color of formaldehyde fixed specimens over time (Kaiserling, 1900; Ajileye and Adeyemi, 2020). Although Kaiserling III is not a pure glycerol solution, as it also contains potassium acetate and chloral hydrate, the state of tissues preserved in it can act as a proxy for evaluating glycerol as a preservative agent. Although Kaiserling originally recommended 25 % glycerol, today, 40 % glycerol has been reported as a better concentration for fluid preservation (Ajileye and Adeyemi, 2020). One of Kaiserling's contributions was identifying the need to minimize air exposure and acid formation within the preservation medium to the greatest extent feasible (Potaliya and Ghatak, 2016). One study evaluated the histologic quality of fixed brain tissue that had been preserved in Kaiserling III for up to 55 years (Monteiro, 2008). When an archival specimen was pre-treated in 10 % formalin prior to histologic processing, it had similar histology as control brain tissue, including intact cellular structure, except for having less distinct nucleoli. Immunostaining was also possible, with neurofilament staining allowing the identification of individual neuronal processes and GFAP staining allowing for the identification of astrocytes. Glycerol has even been used to preserve biospecimens for extended periods without fixatives. For example, skin tissue is often preserved in skin banks using high concentrations of glycerol, up to 98 %. This leads a loss of cellular viability but retention of ultrastructure for years (de Backere, 1994; Richters et al., 1996). Dura mater has been preserved for up to 7 weeks in 98 % glycerol, allowing for the retention of normal ultrastructure, with the exception of more finely divided collagen fibers (McGarvey et al., 1984). Nerve tissue has been preserved in 98 % glycerol at 4 °C for months prior to transplant (Wolff et al., 1993). Regarding brain tissue, one study found that samples of brain tissue can be stored in phosphate-buffered 50 % glycerol at temperatures of 4 °C or at –20 °C, for at least 15 months (Lembo et al., 2006). This storage procedure allowed the tissue to be successfully immunostained for rabies antigen against a neuronal background. Rather than suggesting that glycerol be used as a solitary preservative in brain banking, these studies show the power of glycerol preservation and why combining it with the technique of fixation can be helpful. One downside of glycerol preservation is that, at intermediate concentrations of glycerol of less than 65 % and relative humidities of greater than 75 % in the room used for storage, specimens can be susceptible to mold growth (Van Dam, 2020). A small concentration of fixative, such as 0.5 % formaldehyde, or microbial inhibitors such as camphor or thymol can be added to prevent mold growth (Ajileye and Adeyemi, 2020). Glycerol is also expected to eventually break down into reactive impurities during the storage process, such as via oxidation to methylglyoxal (Sugiura et al., 2020). Finally, storage in glycerol may promote chemical reactions, such as the Maillard reaction, which occurs between amino acids and reducing sugars (Smarrito-Menozzi et al., 2013). Other cryoprotectants Ethylene glycol has also been used as a fluid preservative, most commonly for marine species (Williamson and Russell, 1965). Compared to glycerol, ethylene glycol has a lower risk of causing osmotic damage, allowing for stronger solutions to be used (Williamson and Russell, 1965). However, one source reports that it can cause precipitates over time when used as a fluid preservative (Luisa, 1982, p. 33). Disaccharides are also potential alternatives to glycerol for storage in the liquid state. Trehalose has been found to stabilize biomolecules such as proteins when dissolved in aqueous solutions, potentially due to its effect of raising the surface tension of water (Kaushik and Bhat, 2003). Another source compares heart valve storage in 50-80 % sucrose to pure glycerol at 4 °C, finding that the sucrose solution was able to maintain structural preservation for 52 weeks while glycerol was able to maintain preservation for 12 weeks (as cited in Vásquez-Rivera et al., 2018). When stored in sucrose, some sources recommend to also add sodium azide (Strnad et al., 2022). Kinetics of cryoprotectant brain penetration Many of these studies are performed on small biopsy samples or small organisms. However, it is often desired for the human brain to be stored intact, for example for the purpose of ex vivo neuroimaging, or so that long-range neural connections can be mapped. In this case, there is a need to implement a protocol allowing for the cryoprotectant to penetrate throughout the brain, without taking too long or causing damage due to rapid changes in osmotic concentration. One study performed an immersion of 10 % glycerol and 2 % DMSO into one hemisphere of a fixed rhesus monkey brain for 1 day, followed by immersion in 20 % glycerol and 2 % DMSO for 3 days (Sloane et al., 2000). However, the rhesus brain volume is about 10-15 times less than the human brain volume, meaning that this process may be time consuming in a human brain banking setting. Notably, evidence suggests that osmotic damage is not as significant in tissue that has been adequately fixed (Paljärvi et al., 1979). One alternative possibility would be to perfuse the preservative chemicals, but that is challenging to perform consistently in a postmortem setting, especially because it would need to be done evenly to avoid inducing osmotic damage (McFadden et al., 2019). Summary Cryoprotectants such as glycerol, ethylene glycol, and sucrose have shown promise for room temperature storage of fixed brain tissue while minimizing biomolecular extraction. Glycerol has been used since the late 1800s, with studies generally reporting intact morphology after years or even decades of storage, except for a loss of nucleolar or collagen fiber detail. However, there are not many studies on this topic, and optimal protocols for penetrating large intact human brains have not been established. Overall, cryoprotectants are a promising fluid storage approach in combination with an initial fixation step, warranting further investigation. Other storage conditions Temperature Temperature is a critical factor influencing the rate of chemical reactions during fluid preservation. Lower temperatures slow reaction kinetics, enabling longer preservation durations. As a rule of thumb, the 10-degree rule estimates that chemical and biological reaction rates decrease 2 fold per 10 °C temperature drop (Hukins et al., 2008; Orlowski et al., 2014). Based on this estimate, a brain stored at a standard refrigerator of 4 °C would be expected to have a compared to have a 4-fold slower decomposition rate than one stored at a room temperature of approximately 24 °C. In brain banking, it is common but certainly not ubiquitous to store fixed tissue at refrigerator temperature of around 4 °C to slow degradation (Insausti et al., 1995; Wiggermann et al., 2023). One study reported that storage at 4 °C, alongside a substantially lower concentration of fixative, led to a lesser degree of antigen degradation over time for several antigens compared to storage at higher temperatures and a higher concentration of fixative (Lyck et al., 2008). For alcohol preserved specimens, storing them below the flash point of absolute ethanol of 12 °C is often recommended for safety reasons. However, low temperatures can introduce new problems. Storage below 0 °C risks ice crystal formation and associated tissue damage, unless cryoprotectants are used. Even above 0 °C, lower temperature storage favors the precipitation of polyoxymethylene from formaldehyde (Howe et al., 1995). The main downside of lower temperature storage is that it can substantially increase the costs due to the need for storage infrastructure and labor for maintenance. As a result, despite the expectation that it will slow degradation, storage at low temperature needs to be balanced by cost-benefit considerations. Storage container The container used for fluid preservation plays an important role in preventing evaporation and oxidation, which can lead to severe tissue damage over time. Glass and plastic are the most common materials used. Glass is impermeable to oxygen and moisture but is vulnerable to cracking over time, especially if stressed (Stoddart, 1989; Hiebert et al., 2021). Plastic containers vary in their oxygen and moisture permeability. In long-term fluid preservation, one of the major risks is evaporation leading to tissue desiccation and potential irreversible damage (Eichhorn et al., 2018). Any compromise in the container, such as fissures in the glass, can result in evaporation and specimen desiccation (Williamson and Russell, 1965). For both glass and plastic containers, the integrity of the seal is also critical in minimizing oxygen permeability. Preventing oxygen exposure is essential because oxygen causes oxidative damage to brain tissue and is associated with decreases in pH. Oxidation affects unsaturated linkages in lipids, certain amino acids in proteins, and pigments (Stoddart, 1989). To minimize oxygen exposure, the container can be filled with fluid or otherwise degassed (Stoddart, 1989). If a specimen is stored in an oxygen-sealed container, then every time it is opened, the fluid preservative and specimen will be re-exposed to oxygen (Barnes et al., 2000). As a space-saving measure, some sources recommend storing tissue in heat-sealed plastic bags, but this risks desiccation and oxidative damage over time (Høyer et al., 1991). Adding antioxidants to the preservative solution is another approach to mitigate oxidative damage (Silvestre et al., 2021; Sugiura et al., 2020). Another parameter that containers affect is the amount of light exposure. Minimizing exposure to light is often recommended in biospecimen storage (Maki et al., 2021). UV radiation can cause photodamage to biomolecules via covalent bond formation, such as thymidine dimer formation in DNA (Kansagara et al., 2008). Exposure to light can also cause pigment fading (Monteiro, 2008). Storing specimens in UV-protective containers or avoiding light exposure is recommended. Overall, the container plays a crucial role in fluid preservation by regulating evaporation, oxidation, and light exposure. Fluid refreshing Brain banking protocols often include steps to periodically refresh the preservative fluid. When using formaldehyde solutions, this has be used in an attempt to prevent a decline in pH (Rahimi et al., 2006). For example, multiple sources report refreshing formaldehyde solutions every 2 years (Rahimi et al., 2006; Qiu et al., 2019). However, over time, the rate of biomolecular extraction decreases as the fluid preservative reaches equilibrium with the biospecimen (Simmons, 2019). As a result, a key downside of fluid refreshing is that it will disrupt this equilibrium. Additionally, fluid refreshing will lead to the loss of any small molecules that have leached out into the solution. This is why one source recommends that cocaine levels should be tested in the formalin solution, not just in the tissue (Hilal et al., 2009). Because different tissue types, containers, and fluid compositions will all lead to degradation at different rates, any fluid change schedule should be adapted to the needs and circumstances of the brain bank. More durable containers and fluids will need less frequent refreshing. The optimal schedule of fluid refreshing, if it is netbeneficial at all for expected future research applications, is unknown. Summary Temperature significantly influences the rate of chemical reactions in fluid preservation, with lower temperatures slowing down these reactions. The type of storage container is crucial for preventing tissue damage due to evaporation and oxidation. Glass and plastic are common materials, but their integrity and sealing are vital. Periodic fluid refreshing is sometimes performed in brain banking, often aiming to minimize the accumulation of acidity. However, it can disrupt the brain tissue’s chemical equilibrium and may result in the loss of certain molecules from the solvent. Comparison to alternatives Cryopreservation Cryopreservation uses subzero temperatures to decrease the rate of molecular motion and thereby improve long-term preservation outcomes. Cryopreservation can be performed either without fixation or following fixation, thereby either avoiding entirely or substantially mitigating the problem of overfixation occurring in fluid preservation. Consistent with this, data suggests that the imaging signal for proteins and nucleic acids is improved when comparing cryopreservation to long-term fixation protocols (Kingsbury et al., 1999; Hrabovszky et al., 2007). When used without fixation, cryopreservation also preserves more enzyme activity and allows for the use of many biomolecular profiling techniques that are more difficult to perform on fixed tissue (Verhaert et al., 1990). However, cryopreservation introduces the issue of ice artifact, which can severely damage brain cell morphology (Itoyama et al., 1980; Vonsattel et al., 2008). Ice artifacts can be mitigated or prevented by cryopreserving small tissue segments at a fast rate or by using cryoprotectants. Indeed, fixed and cryoprotected tissue allows for high-quality morphologic preservation (Sloane et al., 2000; Van Herp et al., 2005; Estrada et al., 2017). After long-term storage at moderate subzero temperatures of approximately -20 °C, fixed and sufficiently cryoprotected tissue is expected to be in the fluid state, yet these protocols have shown good preservation of morphology and antigenicity for years (Watson et al., 1986; Otubo et al., 2021). However, cryoprotection in the absence of fixation is challenging, because cryoprotectant immersion requires a slow ramp-up to prevent osmotic damage, and perfusion quality is often limited in the postmortem setting (McFadden et al., 2019). One source reports that even when the cryoprotectant immersion procedure was performed carefully, some freezing damage still occurred; however, the cryoprotectant solutions they used were likely not at the concentration required to vitrify (Romijn et al., 1999). Cryopreservation of large tissue blocks or the entire brain also poses the risk of thermal stress fracturing and physical aging (Chang and Baust, 1991; Gangwar et al., 2022). Perhaps most importantly, cryopreservation is substantially more expensive and less reliable than storage via fluid preservation (Dwork et al., 1998; Leboeuf et al., 2008). Paraffin embedding Embedding specimens in paraffin wax was first introduced in the 1860s (van der Lem et al., 2021). Following the introduction of formaldehyde fixation in the 1890s, formaldehyde fixed paraffin embedded (FFPE) tissue blocks have grown to become one of the most common methods for preserving biospecimens. Paraffin embedding involves multiple steps that can cause damage, including heating and dehydration. Paraffin is made up of a mixture of straight chain alkanes. When the tissue cools, the paraffin crystallizes, allowing the tissue to be stored in the solid state (Rhodes et al., 1927; Zocher and Machado, 1959). Antigenicity preservation can be improved in FFPE by removal of residual water and storing tissue in low humidity environments (Xie et al., 2011). FFPE tissue is very commonly used in archival settings, with demonstrated preservation of biological structures for decades (Bradl and Lassmann, 2012; Rauch et al., 2018; Guerra et al., 2020). Numerous studies have shown that FFPE demonstrates better antigenicity than tissue stored long-term in formaldehyde solutions, consistent with its ability to prevent overfixation (Sillevis Smitt et al., 1993; Dwork et al., 1998; Lin et al., 2023). Also, the localized areas of empty neuropil seen in the one study of brains stored in formalin that identified this artifact were not seen in matched FFPE samples from the same cohort (van Duijn et al., 2011). However, there are drawbacks to paraffin embedding. First, the tissue processing required, especially dehydration, can lead to the loss of numerous biomolecules that are not sufficiently crosslinked, including structural lipids and amino acids (Vos et al., 2019; Dannhorn et al., 2022). While some lipid species are solvent-resistant, possibly due to direct formaldehyde cross-linking, the overall biochemical landscape is irreversibly altered by dehydration (Denti et al., 2022). Additionally, it is not yet possible to perform paraffin embedding on whole human brain samples en bloc. To our knowledge, the largest specimens that have been shown to be embedded in paraffin are less than 60 cm3, such as a macaque brain hemisphere (Zhanmu et al., 2020). This means that cutting damage will occur during to dissection and there will be constraints on the size, type, and orientation of the tissue preserved (Dwork et al., 1998). Finally, paraffin embedding is more complex and expensive than simply allowing the brain to remain in fluid preservative for the long term. Resin embedding Embedding resins such as epoxies and acrylates into brain tissue is commonly used as sample preparation for microscopy studies. As with paraffin embedding, this allows for storage in a solid state, which minimizes overfixation and chemical reactivity over time. However, also as with paraffin embedding, the extensive dehydration required for resin embedding often leads to loss of lipids and other biomolecules that are not directly crosslinked following fixation. The extent of dehydration required varies based on the resin, with some acrylates able to dissolve in up to 10-12 % of water by weight (Newman and Hobot, 1999). The extent of lipid extraction also depends on the resin. For example, the acrylate resin Lowicryl HM20 extracts fewer lipids, because it is nonpolar (Glauert and Lewis, 1998). As they are used for electron microscopy, resin embedding clearly offers excellent morphological preservation. The stability of resin embedded specimens over time is not well established in the literature. There are some theoretical reasons to expect that preservation durations may not be unlimited in the very long term. For example, polymerized epoxy resins are found to act in some circumstances as viscous fluids, allowing for displacement of tissue elements over time (Mollenhauer, 1993). However, most sources suggest that resin embedded tissue has good long-term storage longevity, especially when stored in a desiccator (Glauert and Lewis, 1998, p. 276; Celis, 2006, p. 294; Becker et al., 2014). Resin embedding is often only practical on small tissue samples and the chemicals required can be relatively expensive, proprietary, and challenging to use properly, limiting their widespread use in brain banking. Summary Compared to fluid preservation of brain tissue, cryopreservation offers better antigen preservation and biomolecular profiling capabilities, but faces the downsides of ice artifact and high costs. Paraffin embedding is reliable for long-term storage and antigenicity but leads to the loss of a large subset of biomolecules. Resin embedding is best for morphological preservation but is limited by sample volume and expense. Most brain banks utilize a multifaceted strategy, storing some tissue samples with each of fluid preservation, cryopreservation, and paraffin embedding. This allows brain banks to capitalize on the unique advantages of each method and provide a versatile resource for a range of future experimental needs. Relationship with the post-mortem interval In addition to storage time, another key technical variable in brain banking is the postmortem interval, i.e. the amount of time between when death occurs and when the preservation procedure begins. There are several histologic artifacts known to manifest during the postmortem interval, such as pericellular rarefaction, vacuolization, and cell death due to oncotic necrosis (Krassner et al., 2023). In general, these are distinct from the storage artifacts that we identified as potentially associated with fluid preservation. An important exception is decreased antigenicity on immunohistochemistry, which is a multifactorial process that can be caused by both decomposition associated with the postmortem interval and overfixation due to long periods of preservation in fixative-containing solutions. Rapid fixation approaches such as perfusion fixation or ventricular injection may be able to help ameliorate damage associated with the postmortem interval, by minimizing any additional degradation that occurs prior to the fixative reaching that area of the tissue (McFadden et al., 2019; McKenzie et al., 2022). However, we are not aware of any substantial evidence suggesting that different long-term fluid preservation options may interact with the postmortem interval in a similar way. Notably, for many applications, the age of the fluid-preserved tissue, even after many years, is generally considered to be less impactful on research outcomes than the postmortem interval (Wu et al., 2002; Herbin et al., 2021). Implications for brain banking Investigators operating brain banks and smaller brain collections face many constraints, including limited time and resources. The notion of the “shelf life” of brain tissue is important in brain banking (Erslev, 2018). Our review suggests that fluid-preserved brain tissue can be used in research applications for at least several decades, depending on the research goals. For antigen preservation, long-term immersion in formaldehyde solutions clearly results in overfixation and diminished antigenicity for immunohistochemical studies (Liu et al., 2010). Although contemporary antigen retrieval techniques can mitigate some of the effects of overfixation, they cannot always fully restore the original antigenic properties (Ramos-Vara and Miller, 2014). Therefore, recording and communicating the fixation time is crucial for researchers who will use the tissue. For morphological preservation, the existing literature presents limitations in drawing conclusions. As a result, it is still unclear whether brains stored for long periods of time in fluid preservative will be appropriate for many types of brain mapping study, and this will require further experimental research. Most of the extant histology data is available for long-term storage in formaldehyde solutions. There is relatively less of a database to recommend the use of alternatives such as buffer storage or storage in cryoprotectants. There are also many unknowns in the field, such as the longevity of antimicrobial activity conferred by sodium azide in solution. As a result, there is currently no universally accepted “gold standard” method or performance metrics for fluid preservation in brain banking, with trade-offs among existing preservation methods (Table 1). Regardless of the preservation fluid used, maintaining a low temperature likely improves preservation outcomes. For example, morphological preservation has been reported to be intact after 92 years of storage in formalin at room temperature (Herbin et al., 2021). Based on an assumed Q10 value of 2, this implies a refrigerator temperature storage length of approximately 4 times longer, or 368 years. If refrigeration is not feasible, a cool room with stable temperatures is recommended. Finally, a sturdy container that reduces the risks of evaporation, desiccation, oxidative damage, and light exposure can further enhance tissue quality over the long-term.
Areas for further research One high-priority concern is better characterization of storage artifacts that could impact both short-term and long-term research objectives. Critical among these are the areas of empty spaces in neuropil, whose prevalence and impact on neural circuit mapping are yet to be thoroughly examined (van Duijn et al., 2011). It remains an open question whether the type of container used is a factor in accelerating or mediating this artifact. Given that these areas of empty space in neuropil are associated with white discolorations observable at the macroscopic scale, initial investigations into this artifact could be conducted in different cohorts in a relatively straightforward manner. Advanced imaging techniques like MRI, along with correlative histological analyses, could offer another way to identify these artifacts and any related ones. Another important area is to research why nuclei, nucleoli, and DNA have been found to be vulnerable to alterations during storage in some studies but not others (Cook et al., 2014; Monteiro, 2008; Herbin et al., 2021; Berrino et al., 2022). The stability of other biomolecules, especially lipids, also requires further study. Further research could identify which lipid species are extracted, modified, or cross-linked during storage, and what implications these changes have for research using banked brains. Finally, research comparing the extent of storage artifacts when using different fluid preservation methods would be a valuable contribution to the field. Comparison to other reviews As far as we know, no previous review has solely focused on fluid preservation in brain banking, although several sources have discussed it as a part of a broader focus. One source performs a general review of storage of biospecimens in fluid preservatives (Simmons, 2019). They have a thorough discussion of container choice and note that the storage environment is critical, including temperature control, humidity control, minimizing light and UV radiation exposure, minimizing vibrations, preventing evaporation, and preventing physical damage to the specimen. One review focuses on room temperature storage methods for biospecimens, emphasizing the space, cost, and environmental downsides of low temperature storage (Lou et al., 2014). One review notes that for developmental neurotoxicity testing, nervous system specimens are recommended to be embedded in paraffin or resin for long-term storage, in order to avoid shrinkage artifacts potentially associated with prolonged storage in fixative (Garman et al., 2016). One study searched PubMed and found six studies evaluating the effect of prolonged fixation on neural and glial cell marker antigenicity, covering a partially overlapping literature when compared with our review (Wu et al., 2022). They found that there were clear time- and antigen-dependent effects of prolonged fixation. Strengths and limitations of this review One strength of this review is that we have included a large set of studies, allowing for findings from many years ago to be contextualized with current paradigms. Also, we explored results using both biomolecular profiling and histology, highlighting the connections between these two linked fields. This review also has several limitations. First, we did not review in-depth the data from non-brain organs, even though the results for other organs are likely to be relevant to those from the brain. Second, the included studies usually have qualitative assessments of morphologic preservation, not quantitative ones, which means that subtle shifts in neural circuitry during long-term storage might not be detected. On a related note, studies may be susceptible to sampling bias in the choice of brain tissue they profiled, which is more challenging to address when analyzing qualitative metrics of preservation. Third, we did not perform quality ratings of individual studies, so it is more difficult to appropriately weight the findings of individual studies based on the quality of their evidence. However, with such a heterogenous, small field, it is not clear how to rate the quality of studies. Finally, our search strategy was not exhaustive and likely did not identify all the relevant studies on this topic. However, we believe the studies we identified are likely representative of the broader literature. Conclusions Archival brain tissue is a valuable resource, often providing well-characterized specimens that may be more useful for certain studies than freshly acquired samples (Bradl and Lassmann, 2012). Formaldehyde is the most comprehensively studied preservative and has been found capable of maintaining the intricacies of neural circuitry, such as dendritic protrusions, even over extended storage durations. However, long-term storage in formaldehyde causes overfixation, making tissue difficult to process and hindering antigenicity. Additionally, there appears to be extraction of some biomolecules that are not directly crosslinked, including a subset of lipids and small molecules. Other artifacts have also been suggested to occur in formaldehyde solutions that may be specific to the method of storage or imaging employed. In contrast, alternative fluid preservation solutions, such as cryoprotectants and buffers with sodium azide, could avoid the pitfalls associated with storage in formaldehyde. Simply using a lower concentration of formaldehyde for storage may also help to mitigate these problems. A lack of solid data on these alternative methods, however, means that they cannot yet be recommended as obviously superior. In short, the drawbacks of long-term formaldehyde storage are largely known but not yet fully quantified, and whether alternatives carry their own hidden issues remains an open question in need of further research. Abbreviations BA - Brodmann area, CAMKIIA - Ca2+/calmodulin-dependent protein kinase II a isoform, CLARITY - Clear lipid-exchanged acrylamide-hybridized rigid imaging/immunostaining compatible tissue hydrogel, CUBIC - Clear unobstructed brain/body imaging cocktails and computational analysis, DCX - Doublecortin, d - Day, DMSO - Dimethyl sulfoxide, FFPE - Formaldehyde fixed paraffin embedded, GFAP - Glial fibrillary acidic protein, ISH - In situ hybridization, MRI - Magnetic resonance imaging, miRNA - Micro RNA, n - Sample size, NBF - Neutral buffered formalin, NeuN - Neuronal nuclear protein, PSA-NCAM - Polysialylated-neural cell adhesion molecule, PSD95 - Postsynaptic density protein 95, Q10 - Temperature coefficient, RAMESES - Realist and meta-narrative evidence syntheses evolving standards, sDISCO - Stabilized 3D imaging of solvent cleared organs, SHANEL - Small-micelle-mediated human organ efficient clearing and labeling, SMI312 - A commercial antibody staining for neurofilaments from Sternberger Monoclonals Incorporated, SWITCH - System-wide control of interaction time and kinetics of chemicals, vGlut1 - Vesicular glutamate transporter 1, ZO-1 - Zonula Occludens-1. Author contributions A.M., O.N., K.F., and J.C. conceptualized the study. A.M. performed abstract and article screening. A.M. performed data extraction from the included studies and O.N. reviewed the data extracted from a subset of the studies. A.M. and O.N. performed grading of the storage artifacts. K.S. and E.T. performed imaging of the gross brain specimens and associated data analysis. A.M. wrote the initial draft of the manuscript. All authors reviewed the manuscript and approved the final manuscript. Acknowledgements We would like to thank Louise van der Weerd, Andrew Dwork, Thomas Beach, Patrick Hof, Zhuhao Wu, John Smart, Kenneth Hayworth, Martin Baldan, Trevor Christensen, Aschwin de Wolf, Aurelia Song, Anna LaVergne, and Borys Wrobel for helpful personal communications regarding this topic. The Icahn School of Medicine at Mount Sinai provided access to library resources. Funding The study was supported by NIH grants P30AG066514, RF1MH128969, R01AG054008, RF1NS095252, RF1AG060961, R01NS086736, R01AG062348, K01AG070326, and the Rainwater Charitable Foundation. The funders had no role in the design of the study or in the collection or interpretation of the data. Conflict of interest Andrew McKenzie is employed by Oregon Brain Preservation, a non-profit brain preservation organization. The management of Oregon Brain Preservation had no role in the design of the study or in the collection or interpretation of the data. Data availability The generated database of studies and extracted data is available in table format at https://github.com/andymckenzie/Fluid_Preservation This repository also contains the code required to reproduce the figure created in R. Supplementary material Supplementary Material 1 (download) - Rameses checklist (DOCX) Supplementary Material 2 (download) - Additional review methods (DOCX) Supplementary Material 3 (download) - List of PubMed IDs from initial literature search (TXT) Supplementary Material 4 (download) - Articles selected after abstract screening (XML/zip) Supplementary Material 5 (download) - Articles selected after full text review (XML/zip) Supplementary Material 6 (download) - Information about the included studies (PDF) Supplementary Material 7 (download) - Extracted data from the studies using aldehyde storage (PDF) Supplementary Material 8 (download) - Extracted data from the studies using buffer storage (PDF) Supplementary Material 9 (download) - Extracted data from the studies using alcohol storage (PDF) Supplementary Material 10 (download) - Extracted data from the studies using tissue cleared stoarge (PDF) Supplementary Material 11 (download) - Extracted data from the studies using cryoprotectant storage (PDF) References Adams, D.J., 2022. Personal Perspective on Understanding Low Molecular Weight Gels. J. Am. Chem. Soc. 144, 11047–11053. https://doi.org/10.1021/jacs.2c02096 Ajileye, A., Adeyemi, O., 2020. Museum Techniques and Practices in Histopathology: A Compendium. J. Med. Lab. Sci. 30, 18–34. https://doi.org/10.5281/zenodo.4038511 Arends, F., Nowald, C., Pflieger, K., Boettcher, K., Zahler, S., Lieleg, O., 2015. The biophysical properties of Basal lamina gels depend on the biochemical composition of the gel. PloS One 10, e0118090. https://doi.org/10.1371/journal.pone.0118090 Argüelles-Monal, W., Goycoolea, F.M., Peniche, C., Higuera-Ciapara, I., 1998. Rheological study of the chitosan/glutaraldehyde chemical gel system. Polym. Gels Netw. 6, 429–440. https://doi.org/10.1016/S0966-7822(98)00032-X Armitage, W.J., 1986. Osmotic stress as a factor in the detrimental effect of glycerol on human platelets. Cryobiology 23, 116–125. https://doi.org/10.1016/0011-2240(86)90002-7 Bahr, G.F., Bloom, G., Friberg, U., 1957. Volume changes of tissues in physiological fluids during fixation in osmium tetroxide or formaldehyde and during subsequent treatment. Exp. Cell Res. 12, 342–355. https://doi.org/10.1016/0014-4827(57)90148-9 Baird, S.F., 1859. Directions for collecting, preserving and transporting specimens of natural history. Smithsonian institution. Barker, H., Oliver, R., Grant, R., Stephen, L., 1980. Formaldehyde as a pre-treatment for dermal collagen heterografts. Biochim. Biophys. Acta 632, 589–597. https://doi.org/10.1016/0304-4165(80)90335-9 Barnes, I., Holton, J., Vaira, D., Spigelman, M., Thomas, M.G., 2000. An assessment of the long-term preservation of the DNA of a bacterial pathogen in ethanol-preserved archival material. J. Pathol. 192, 554–559. https://doi.org/10.1002/1096-9896(2000)9999:9999<::AID-PATH768>3.0.CO;2-C Baydin, T., Aarstad, O.A., Dille, M.J., Hattrem, M.N., Draget, K.I., 2022. Long-term storage stability of type A and type B gelatin gels: The effect of Bloom strength and co-solutes. Food Hydrocoll. 127, 107535. https://doi.org/10.1016/j.foodhyd.2022.107535 Beach, T.G., Adler, C.H., Sue, L.I., Serrano, G., Shill, H.A., Walker, D.G., Lue, L., Roher, A.E., Dugger, B.N., Maarouf, C., Birdsill, A.C., Intorcia, A., Saxon-Labelle, M., Pullen, J., Scroggins, A., Filon, J., Scott, S., Hoffman, B., Garcia, A., Caviness, J.N., Hentz, J.G., Driver-Dunckley, E., Jacobson, S.A., Davis, K.J., Belden, C.M., Long, K.E., Malek-Ahmadi, M., Powell, J.J., Gale, L.D., Nicholson, L.R., Caselli, R.J., Woodruff, B.K., Rapscak, S.Z., Ahern, G.L., Shi, J., Burke, A.D., Reiman, E.M., Sabbagh, M.N., 2015. Arizona study of aging and neurodegenerative disorders and brain and body donation program. Neuropathology 35, 354–389. https://doi.org/10.1111/neup.12189 Beach, T.G., Tago, H., Nagai, T., Kimura, H., McGeer, P.L., McGeer, E.G., 1987. Perfusion-fixation of the human brain for immunohistochemistry: comparison with immersion-fixation. J. Neurosci. Methods 19, 183–192. https://doi.org/10.1016/s0165-0270(87)80001-8 Becker, K., Hahn, C.M., Saghafi, S., Jährling, N., Wanis, M., Dodt, H.-U., 2014. Reduction of photo bleaching and long term archiving of chemically cleared GFP-expressing mouse brains. PloS One 9, e114149. https://doi.org/10.1371/journal.pone.0114149 Becker, K., Jährling, N., Saghafi, S., Weiler, R., Dodt, H.-U., 2012. Chemical clearing and dehydration of GFP expressing mouse brains. PloS One 7, e33916. https://doi.org/10.1371/journal.pone.0033916 Beckstead, J.H., 1994. A simple technique for preservation of fixation-sensitive antigens in paraffin-embedded tissues. J. Histochem. Cytochem. Off. J. Histochem. Soc. 42, 1127–1134. https://doi.org/10.1177/42.8.8027531 Bekkouche, B.M.B., Fritz, H.K.M., Rigosi, E., O’Carroll, D.C., 2020. Comparison of Transparency and Shrinkage During Clearing of Insect Brains Using Media With Tunable Refractive Index. Front. Neuroanat. 14, 599282. https://doi.org/10.3389/fnana.2020.599282 Berrino, E., Annaratone, L., Detillo, P., Grassini, D., Bragoni, A., Sapino, A., Bussolati, B., Bussolati, G., Marchiò, C., 2022. Tissue Fixation with a Formic Acid-Deprived Formalin Better Preserves DNA Integrity over Time. Pathobiol. J. Immunopathol. Mol. Cell. Biol. 1–11. https://doi.org/10.1159/000525523 Bhatia, H.S., Brunner, A.-D., Öztürk, F., Kapoor, S., Rong, Z., Mai, H., Thielert, M., Ali, M., Al-Maskari, R., Paetzold, J.C., Kofler, F., Todorov, M.I., Molbay, M., Kolabas, Z.I., Negwer, M., Hoeher, L., Steinke, H., Dima, A., Gupta, B., Kaltenecker, D., Caliskan, Ö.S., Brandt, D., Krahmer, N., Müller, S., Lichtenthaler, S.F., Hellal, F., Bechmann, I., Menze, B., Theis, F., Mann, M., Ertürk, A., 2022. Spatial proteomics in three-dimensional intact specimens. Cell 185, 5040-5058.e19. https://doi.org/10.1016/j.cell.2022.11.021 Bien, T., Bessler, S., Dreisewerd, K., Soltwisch, J., 2021. Transmission-Mode MALDI Mass Spectrometry Imaging of Single Cells: Optimizing Sample Preparation Protocols. Anal. Chem. 93, 4513–4520. https://doi.org/10.1021/acs.analchem.0c04905 Boucher, R.M., 1978. Stability of glutaraldehyde disinfectants during storage and use in hospitals. Respir. Care 23, 1063–1072. Bouvier, D.S., Jones, E.V., Quesseveur, G., Davoli, M.A., A Ferreira, T., Quirion, R., Mechawar, N., Murai, K.K., 2016. High Resolution Dissection of Reactive Glial Nets in Alzheimer’s Disease. Sci. Rep. 6, 24544. https://doi.org/10.1038/srep24544 Boyer, I.J., Heldreth, B., Bergfeld, W.F., Belsito, D.V., Hill, R.A., Klaassen, C.D., Liebler, D.C., Marks, J.G., Shank, R.C., Slaga, T.J., Snyder, P.W., Andersen, F.A., 2013. Amended Safety Assessment of Formaldehyde and Methylene Glycol as Used in Cosmetics. Int. J. Toxicol. 32, 5S-32S. https://doi.org/10.1177/1091581813511831 Bradl, M., Lassmann, H., 2012. Microarray analysis on archival multiple sclerosis tissue: pathogenic authenticity outweighs technical obstacles. Neuropathol. Off. J. Jpn. Soc. Neuropathol. 32, 463–466. https://doi.org/10.1111/j.1440-1789.2011.01282.x Brown, P., Wolff, A., Gajdusek, D.C., 1990. A simple and effective method for inactivating virus infectivity in formalin-fixed tissue samples from patients with Creutzfeldt-Jakob disease. Neurology 40, 887–890. https://doi.org/10.1212/wnl.40.6.887 Carlos, A.F., Poloni, T.E., Medici, V., Chikhladze, M., Guaita, A., Ceroni, M., 2019. From brain collections to modern brain banks: A historical perspective. Alzheimers Dement. Transl. Res. Clin. Interv. 5, 52–60. https://doi.org/10.1016/j.trci.2018.12.002 Carter, C.L., Jones, J.W., Farese, A.M., MacVittie, T.J., Kane, M.A., 2016. Inflation-Fixation Method for Lipidomic Mapping of Lung Biopsies by Matrix Assisted Laser Desorption/Ionization-Mass Spectrometry Imaging. Anal. Chem. 88, 4788–4794. https://doi.org/10.1021/acs.analchem.6b00165 Carter, C.L., McLeod, C.W., Bunch, J., 2011. Imaging of phospholipids in formalin fixed rat brain sections by matrix assisted laser desorption/ionization mass spectrometry. J. Am. Soc. Mass Spectrom. 22, 1991–1998. https://doi.org/10.1007/s13361-011-0227-4 Celis, J.E., 2006. Cell Biology: A Laboratory Handbook. Elsevier. Chang, Z.H., Baust, J.G., 1991. Physical aging of glassy state: DSC study of vitrified glycerol systems. Cryobiology 28, 87–95. https://doi.org/10.1016/0011-2240(91)90010-l Chen, K., Muthukumar, M., 2021. Entropic barrier of topologically immobilized DNA in hydrogels. Proc. Natl. Acad. Sci. U. S. A. 118, e2106380118. https://doi.org/10.1073/pnas.2106380118 Clark, A.H., 1991. Structural and Mechanical Properties of Biopolymer Gels, in: Dickinson, E. (Ed.), Food Polymers, Gels and Colloids. Woodhead Publishing, pp. 322–338. https://doi.org/10.1533/9781845698331.322 Cook, S., Dodge, C., Morgan, R., Sandusky, G.E., 2014. DNA/RNA degradation rate in long term fixed museum specimens. Forensic Med. Anat. Res. 3, 1. https://doi.org/10.4236/fmar.2015.31001 Dannhorn, A., Swales, J.G., Hamm, G., Strittmatter, N., Kudo, H., Maglennon, G., Goodwin, R.J.A., Takats, Z., 2022. Evaluation of Formalin-Fixed and FFPE Tissues for Spatially Resolved Metabolomics and Drug Distribution Studies. Pharmaceuticals 15, 1307. https://doi.org/10.3390/ph15111307 de Backere, A.C., 1994. Euro Skin Bank: large scale skin-banking in Europe based on glycerol-preservation of donor skin. Burns J. Int. Soc. Burn Inj. 20 Suppl 1, S4-9. https://doi.org/10.1016/0305-4179(94)90080-9 de Guzman, A.E., Wong, M.D., Gleave, J.A., Nieman, B.J., 2016. Variations in post-perfusion immersion fixation and storage alter MRI measurements of mouse brain morphometry. NeuroImage 142, 687–695. https://doi.org/10.1016/j.neuroimage.2016.06.028 de Prisco, N., Chemiakine, A., Lee, W., Botta, S., Gennarino, V.A., 2022. Protocol to assess the effect of disease-driving variants on mouse brain morphology and primary hippocampal neurons. STAR Protoc. 3, 101244. https://doi.org/10.1016/j.xpro.2022.101244 Del Signore, F., Arribarat, G., Della Salda, L., Mogicato, G., Deviers, A., Cartiaux, B., Vignoli, M., Peran, P., de Pasquale, F., 2022. Volumetric assessment and longitudinal changes of subcortical structures in formalinized Beagle brains. PloS One 17, e0261484. https://doi.org/10.1371/journal.pone.0261484 Denti, V., Capitoli, G., Piga, I., Clerici, F., Pagani, L., Criscuolo, L., Bindi, G., Principi, L., Chinello, C., Paglia, G., Magni, F., Smith, A., 2022. Spatial Multiomics of Lipids, N-Glycans, and Tryptic Peptides on a Single FFPE Tissue Section. J. Proteome Res. 21, 2798–2809. https://doi.org/10.1021/acs.jproteome.2c00601 Denti, V., Piga, I., Guarnerio, S., Clerici, F., Ivanova, M., Chinello, C., Paglia, G., Magni, F., Smith, A., 2020. Antigen Retrieval and Its Effect on the MALDI-MSI of Lipids in Formalin-Fixed Paraffin-Embedded Tissue. J. Am. Soc. Mass Spectrom. 31, 1619–1624. https://doi.org/10.1021/jasms.0c00208 Douglas, J.F., 2018. Weak and Strong Gels and the Emergence of the Amorphous Solid State. Gels Basel Switz. 4, E19. https://doi.org/10.3390/gels4010019 Dwork, A.J., Liu, D., Kaufman, M.A., Prohovnik, I., 1998. Archival, formalin-fixed tissue: its use in the study of Alzheimer’s type changes. Clin. Neuropathol. 17, 45–49. Dykstra, M., 2010. Why 4F:1G Fixative Works for Me. Microsc. Today 18, 50–53. https://doi.org/10.1017/S1551929510000155 Eberhardt, F., Bushong, E.A., Phan, S., Peltier, S., Monteagudo, P., Weinkauf, T., Herz, A.V.M., Stemmler, M., Ellisman, M., 2022. A uniform and isotropic cytoskeletal tiling fills dendritic spines. eNeuro ENEURO.0342-22.2022. https://doi.org/10.1523/ENEURO.0342-22.2022 Eichhorn, P., Andraschke, U., Dross, F., Geppert, C.I., Hartmann, A., Rau, T.T., 2018. Restoration of an academic historical gross pathology collection-refreshed impact on current medical teaching? Virchows Arch. Int. J. Pathol. 473, 219–228. https://doi.org/10.1007/s00428-018-2369-2 Ericsson, J.L., Saladino, A.J., Trump, B.F., 1965. Electron microscopic observations of the influence of different fixatives on the appearance of cellular ultrastructure. Z. Zellforsch. Mikrosk. Anat. Vienna Austria 1948 66, 161–181. https://doi.org/10.1007/BF00344332 Erslev, T., 2018. A brain worth keeping? Waste, value and time in contemporary brain banking. Stud. Hist. Philos. Biol. Biomed. Sci. 67, 16–23. https://doi.org/10.1016/j.shpsc.2017.12.002 Estrada, L.I., Robinson, A.A., Amaral, A.C., Giannaris, E.L., Heyworth, N.C., Mortazavi, F., Ngwenya, L.B., Roberts, D.E., Cabral, H.J., Killiany, R.J., Rosene, D.L., 2017. Evaluation of long-term cryostorage of brain tissue sections for quantitative histochemistry. J. Histochem. Cytochem. 65, 153–171. https://doi.org/10.1369/0022155416686934 Evers, P., Uylings, H.B., 1997. An optimal antigen retrieval method suitable for different antibodies on human brain tissue stored for several years in formaldehyde fixative. J. Neurosci. Methods 72, 197–207. https://doi.org/10.1016/s0165-0270(96)02204-2 Fahey, S.J., Healy, J.M., 2003. Sperm ultrastructure in the nudibranch genus Halgerda with reference to other Discodorididae and to Chromodorididae (Mollusca: Opisthobranchia). J. Morphol. 257, 9–21. https://doi.org/10.1002/jmor.10086 Fang, S.-G., Wan, Q.-H., Fujihara, N., 2002. Formalin removal from archival tissue by critical point drying. BioTechniques 33, 604, 606, 608–610. https://doi.org/10.2144/02333rr03 Feirabend, H.K., Ploeger, S., 1991. Microwave applications in classical staining methods in formalin-fixed human brain tissue: a comparison between heating with microwave and conventional ovens. Eur. J. Morphol. 29, 185–197. Fish, P.A., 1895. Formalin for the preservation of brains. J. Comp. Neurol. 5, 126–128. https://doi.org/10.1002/cne.910050205 Flor-García, M., Terreros-Roncal, J., Moreno-Jiménez, E.P., Ávila, J., Rábano, A., Llorens-Martín, M., 2020. Unraveling human adult hippocampal neurogenesis. Nat. Protoc. 15, 668–693. https://doi.org/10.1038/s41596-019-0267-y Fox, C.H., Johnson, F.B., Whiting, J., Roller, P.P., 1985. Formaldehyde fixation. J. Histochem. Cytochem. Off. J. Histochem. Soc. 33, 845–853. https://doi.org/10.1177/33.8.3894502 French, D., Edsall, J.T., 1945. The Reactions of Formaldehyde with Amino Acids and Proteins, in: Anson, M.L., Edsall, J.T. (Eds.), Advances in Protein Chemistry. Academic Press, pp. 277–335. https://doi.org/10.1016/S0065-3233(08)60627-0 Furuta, T., Yamauchi, K., Okamoto, S., Takahashi, M., Kakuta, S., Ishida, Y., Takenaka, A., Yoshida, A., Uchiyama, Y., Koike, M., Isa, K., Isa, T., Hioki, H., 2022. Multi-scale light microscopy/electron microscopy neuronal imaging from brain to synapse with a tissue clearing method, ScaleSF. iScience 25, 103601. https://doi.org/10.1016/j.isci.2021.103601 Gangwar, L., Phatak, S.S., Etheridge, M., Bischof, J.C., 2022. A guide to successful mL to L scale vitrification and rewarming. Cryo Letters 43, 316–321. https://doi.org/10.54680/fr22610110112 Garman, R.H., 2011. Common Histological Artifacts in Nervous System Tissues, in: Fundamental Neuropathology for Pathologists and Toxicologists. John Wiley & Sons, Ltd, pp. 191–201. https://doi.org/10.1002/9780470939956.ch13 Garman, R.H., Li, A.A., Kaufmann, W., Auer, R.N., Bolon, B., 2016. Recommended Methods for Brain Processing and Quantitative Analysis in Rodent Developmental Neurotoxicity Studies. Toxicol. Pathol. 44, 14–42. https://doi.org/10.1177/0192623315596858 Gaudin, M., Panchal, M., Ayciriex, S., Werner, E., Brunelle, A., Touboul, D., Boursier-Neyret, C., Auzeil, N., Walther, B., Duyckaerts, C., Laprévote, O., 2014. Ultra performance liquid chromatography - mass spectrometry studies of formalin-induced alterations of human brain lipidome. J. Mass Spectrom. JMS 49, 1035–1042. https://doi.org/10.1002/jms.3424 Gavrilov, A., Razin, S.V., Cavalli, G., 2015. In vivo formaldehyde cross-linking: it is time for black box analysis. Brief. Funct. Genomics 14, 163–165. https://doi.org/10.1093/bfgp/elu037 Gellein, K., Flaten, T.P., Erikson, K.M., Aschner, M., Syversen, T., 2008. Leaching of trace elements from biological tissue by formalin fixation. Biol. Trace Elem. Res. 121, 221–225. https://doi.org/10.1007/s12011-007-8051-1 Glauert, A., Lewis, P., 1998. Biological Specimen Preparation for Transmission Electron Microscopy. Princeton University Press. Grinberg, L.T., Amaro, E., Teipel, S., dos Santos, D.D., Pasqualucci, C.A., Leite, R.E.P., Camargo, C.R., Gonçalves, J.A., Sanches, A.G., Santana, M., Ferretti, R.E.L., Jacob-Filho, W., Nitrini, R., Heinsen, H., Brazilian Aging Brain Study Group, 2008. Assessment of factors that confound MRI and neuropathological correlation of human postmortem brain tissue. Cell Tissue Bank. 9, 195–203. https://doi.org/10.1007/s10561-008-9080-5 Guerra, J.M., Monteiro, R.L., Gonzalez, L., Kimura, L.M., Cirqueira, C.D.S., de Araújo, L.J.T., 2020. Against All Odds: RNA Extraction From Different Protocols Adapted to Formalin-fixed Paraffin-embedded Tissue. Appl. Immunohistochem. Mol. Morphol. AIMM 28, 403–410. https://doi.org/10.1097/PAI.0000000000000772 Gulrez, S.K., Al-Assaf, S., Phillips, G.O., 2011. Hydrogels: methods of preparation, characterisation and applications. Prog. Mol. Environ. Bioeng.- Anal. Model. Technol. Appl. 117150. https://doi.org/10.5772/24553 Guo, Y., Ma, J., Dang, K., Li, Z., Ge, Q., Huang, Y., Wang, G., Zhao, X., 2023. Transcriptomic profiling of nuclei from PFA-fixed and FFPE brain tissues. https://doi.org/10.1101/2023.04.13.536693 Gustafsson, O.J.R., Arentz, G., Hoffmann, P., 2015. Proteomic developments in the analysis of formalin-fixed tissue. Biochim. Biophys. Acta 1854, 559–580. https://doi.org/10.1016/j.bbapap.2014.10.003 Hackett, M.J., McQuillan, J.A., El-Assaad, F., Aitken, J.B., Levina, A., Cohen, D.D., Siegele, R., Carter, E.A., Grau, G.E., Hunt, N.H., Lay, P.A., 2011. Chemical alterations to murine brain tissue induced by formalin fixation: implications for biospectroscopic imaging and mapping studies of disease pathogenesis. The Analyst 136, 2941–2952. https://doi.org/10.1039/c0an00269k Hahn, C., Becker, K., Saghafi, S., Pende, M., Avdibašić, A., Foroughipour, M., Heinz, D.E., Wotjak, C.T., Dodt, H.-U., 2019. High-resolution imaging of fluorescent whole mouse brains using stabilised organic media (sDISCO). J. Biophotonics 12, e201800368. https://doi.org/10.1002/jbio.201800368 Hahn, E.E., Alexander, M.R., Grealy, A., Stiller, J., Gardiner, D.M., Holleley, C.E., 2022. Unlocking inaccessible historical genomes preserved in formalin. Mol. Ecol. Resour. 22, 2130–2147. https://doi.org/10.1111/1755-0998.13505 Hammer, N., Löffler, S., Feja, C., Sandrock, M., Schmidt, W., Bechmann, I., Steinke, H., 2012. Ethanol-glycerin fixation with thymol conservation: a potential alternative to formaldehyde and phenol embalming. Anat. Sci. Educ. 5, 225–233. https://doi.org/10.1002/ase.1270 Hardy, W.B., 1899. On the structure of cell protoplasm: Part I. The Structure produced in a Cell by Fixative and Post-mortem change. The Structure of Colloidal matter and the Mechanism of Setting and of Coagulation. J. Physiol. 24, 158. https://doi.org/10.1113/jphysiol.1899.sp000755 Harris, A., Roseborough, A., Mor, R., Yeung, K.K.-C., Whitehead, S.N., 2020. Ganglioside Detection from Formalin-Fixed Human Brain Tissue Utilizing MALDI Imaging Mass Spectrometry. J. Am. Soc. Mass Spectrom. 31, 479–487. https://doi.org/10.1021/jasms.9b00110 Hartman, T.P.V., 2019. Fluid Preservation of Zoological Specimens. Methods Mol. Biol. Clifton NJ 1897, 135–148. https://doi.org/10.1007/978-1-4939-8935-5_14 Hassani, A., Khan, G., 2015. A simple procedure for the extraction of DNA from long-term formalin-preserved brain tissues for the detection of EBV by PCR. Exp. Mol. Pathol. 99, 558–563. https://doi.org/10.1016/j.yexmp.2015.09.013 Heinsen, H., Arzberger, T., Schmitz, C., 2000. Celloidin mounting (embedding without infiltration) - a new, simple and reliable method for producing serial sections of high thickness through complete human brains and its application to stereological and immunohistochemical investigations. J. Chem. Neuroanat. 20, 49–59. https://doi.org/10.1016/s0891-0618(00)00067-3 Helms, G., Schulz-Schaeffer, W., Schweizer, R., 2014. The preservation state of ethanol-fixated historic brain specimens revealed by quantitative MRI. Presented at the Joint Annual Meeting ISMRM-ESMRMB, Milan, Italy, p. Abstract #1781. Herai, R.R., Stefanacci, L., Hrvoj-Mihic, B., Chailangkarn, T., Hanson, K., Semendeferi, K., Muotri, A.R., 2014. Micro RNA detection in long-term fixed tissue of cortical glutamatergic pyramidal neurons after targeted laser-capture neuroanatomical microdissection. J. Neurosci. Methods 235, 76–82. https://doi.org/10.1016/j.jneumeth.2014.06.028 Herbin, M., Santin, M.D., Toro, R., Heuer, K., Cuisin, J., Martin, C., Raveendran, L., François, C., Debruyne, R., 2021. Do not Dispose of Historic Fluid Collections: Evaluating Research Potential and Range of Use. Collect. Forum 34, 157–169. https://doi.org/10.14351/0831-4985-34.1.157 Heslinga, F.J., Deierkauf, F.A., 1961. The action of histological fixatives on tissue lipids. Comparison of the action of several fixatives using paper chromatography. J. Histochem. Cytochem. Off. J. Histochem. Soc. 9, 572–577. https://doi.org/10.1177/9.5.572 Hiebert, M., Lam, T., Vicenzi, E., Phaneuf, R., Moser, W., Hawks, C., 2021. Microanalysis of Glass Fluid Storage Vials from The Invertebrate Zoology Collection at the National Museum of Natural History. Microsc. Microanal. 27, 3208–3210. https://doi.org/10.1017/S1431927621011077 Hilal, A., Dağlioğlu, N., Battal, D., Yener, F., Dağlioğlu, K., 2009. Detection of cocaine and benzoylecgonine in formalin fixed rat tissues. Toxicol. Mech. Methods 19, 422–426. https://doi.org/10.1080/15376510903147132 Hilton, E.J., Watkins-Colwell, G.J., Huber, S.K., 2021. The expanding role of natural history collections. Ichthyol. Herpetol. 109, 379–391. https://doi.org/10.1643/t2020018 Hoffman, E.A., Frey, B.L., Smith, L.M., Auble, D.T., 2015. Formaldehyde crosslinking: a tool for the study of chromatin complexes. J. Biol. Chem. 290, 26404–26411. https://doi.org/10.1074/jbc.R115.651679 Howe, G.E., Marking, L.L., Bills, T.D., Schreier, T.M., 1995. Efficacy and toxicity of formalin solutions containing paraformaldehyde for fish and egg treatments. Progress. Fish-Cult. 57, 147–152. https://doi.org/10.1577/1548-8640(1995)057<0147:EATOFS>2.3.CO;2 Høyer, P.E., Lyon, H., Møller, M., Prentø, P., van Deurs, B., Hasselager, E., Andersen, A.P., 1991. Tissue Processing: III. Fixation, General Aspects, in: Lyon, Hans (Ed.), Theory and Strategy in Histochemistry: A Guide to the Selection and Understanding of Techniques. Springer, Berlin, Heidelberg, pp. 171–186. https://doi.org/10.1007/978-3-642-73742-8_12 Hrabovszky, E., Kalló, I., Szlávik, N., Keller, E., Merchenthaler, I., Liposits, Z., 2007. Gonadotropin-releasing hormone neurons express estrogen receptor-beta. J. Clin. Endocrinol. Metab. https://doi.org/10.1210/jc.2006-2819 Huang, X., Ahn, D.U., 2019. Lipid oxidation and its implications to meat quality and human health. Food Sci. Biotechnol. 28, 1275–1285. https://doi.org/10.1007/s10068-019-00631-7 Hukins, D.W.L., Mahomed, A., Kukureka, S.N., 2008. Accelerated aging for testing polymeric biomaterials and medical devices. Med. Eng. Phys. 30, 1270–1274. https://doi.org/10.1016/j.medengphy.2008.06.001 Hykin, S.M., Bi, K., McGuire, J.A., 2015. Fixing Formalin: A Method to Recover Genomic-Scale DNA Sequence Data from Formalin-Fixed Museum Specimens Using High-Throughput Sequencing. PloS One 10, e0141579. https://doi.org/10.1371/journal.pone.0141579 Insausti, R., Tuñón, T., Sobreviela, T., Insausti, A.M., Gonzalo, L.M., 1995. The human entorhinal cortex: a cytoarchitectonic analysis. J. Comp. Neurol. 355, 171–198. https://doi.org/10.1002/cne.903550203 Ishii, H., Shirai, T., Makino, C., Nishikata, T., 2014. Mitochondrial inhibitor sodium azide inhibits the reorganization of mitochondria-rich cytoplasm and the establishment of the anteroposterior axis in ascidian embryo. Dev. Growth Differ. 56, 175–188. https://doi.org/10.1111/dgd.12117 Itoyama, Y., Sternberger, N.H., Kies, M.W., Cohen, S.R., Richardson, E.P., Webster, H., 1980. Immunocytochemical method to identify myelin basic protein in oligodendroglia and myelin sheaths of the human nervous system. Ann. Neurol. 7, 157–166. https://doi.org/10.1002/ana.410070211 Ivanchenko, M.V., Indzhykulian, A.A., Corey, D.P., 2021. Electron Microscopy Techniques for Investigating Structure and Composition of Hair-Cell Stereociliary Bundles. Front. Cell Dev. Biol. 9, 744248. https://doi.org/10.3389/fcell.2021.744248 Jia, H., Pu, W.-F., Zhao, J.-Z., Liao, R., 2011. Experimental Investigation of the Novel Phenol−Formaldehyde Cross-Linking HPAM Gel System: Based on the Secondary Cross-Linking Method of Organic Cross-Linkers and Its Gelation Performance Study after Flowing through Porous Media. Energy Fuels 25, 727–736. https://doi.org/10.1021/ef101334y Kaiserling, C., 1900. Über Konservierung und Aufstellung pathologisch-anatomischer Präparate für Schau- und Lehrsammlungen. Verhandl Dtsch. Pathol Gesel 2, 203–217. Kamps, J.J.A.G., Hopkinson, R.J., Schofield, C.J., Claridge, T.D.W., 2019. How formaldehyde reacts with amino acids. Commun. Chem. 2, 1–14. https://doi.org/10.1038/s42004-019-0224-2 Kansagara, A.G., McMahon, H.E., Hogan, M.E., 2008. Dry-state, room-temperature storage of DNA and RNA. Nat. Methods 5. https://doi.org/10.1038/nmeth.f.219 Kaushik, J.K., Bhat, R., 2003. Why is trehalose an exceptional protein stabilizer? An analysis of the thermal stability of proteins in the presence of the compatible osmolyte trehalose. J. Biol. Chem. 278, 26458–26465. https://doi.org/10.1074/jbc.M300815200 Ke, M.-T., Imai, T., 2014. Optical clearing of fixed brain samples using SeeDB. Curr. Protoc. Neurosci. 66, Unit 2.22. https://doi.org/10.1002/0471142301.ns0222s66 Kingsbury, A.E., Mardsen, C.D., Foster, O.J., 1999. DNA fragmentation in human substantia nigra: apoptosis or perimortem effect? Mov. Disord. Off. J. Mov. Disord. Soc. 13, 877–84. https://doi.org/10.1002/mds.870130604 Koehler, J.W., Miller, A.D., Rissi, D.R., 2024. Effects of autolysis and prolonged formalin fixation on histomorphology and immunohistochemistry of normal canine brain tissue: an experimental study. J. Vet. Diagn. Invest. 10406387231220649. https://doi.org/10.1177/10406387231220649 Koo, T.K., Li, M.Y., 2016. A Guideline of Selecting and Reporting Intraclass Correlation Coefficients for Reliability Research. J. Chiropr. Med. 15, 155–163. https://doi.org/10.1016/j.jcm.2016.02.012 Kösel, S., Graeber, M.B., 1994. Use of neuropathological tissue for molecular genetic studies: parameters affecting DNA extraction and polymerase chain reaction. Acta Neuropathol. (Berl.) 88, 19–25. https://doi.org/10.1007/BF00294355 Kotnala, A., Anderson, D.M.G., Patterson, N.H., Cantrell, L.S., Messinger, J.D., Curcio, C.A., Schey, K.L., 2021. Tissue fixation effects on human retinal lipid analysis by MALDI imaging and LC-MS/MS technologies. J. Mass Spectrom. JMS 56, e4798. https://doi.org/10.1002/jms.4798 Kövari, E., Hof, P.R., Bouras, C., 2011. The Geneva brain collection. Ann. N. Y. Acad. Sci. 1225 Suppl 1, E131-146. https://doi.org/10.1111/j.1749-6632.2011.06008.x Kowalczuk, J., Rachocki, A., Bielejewski, M., Tritt-Goc, J., 2016. Effect of gel matrix confinement on the solvent dynamics in supramolecular gels. J. Colloid Interface Sci. 472, 60–68. https://doi.org/10.1016/j.jcis.2016.03.033 Krassner, M.M., Kauffman, J., Sowa, A., Cialowicz, K., Walsh, S., Farrell, K., Crary, J.F., McKenzie, A.T., 2023. Postmortem changes in brain cell structure: a review. Free Neuropathol. 4, 4–10. https://doi.org/10.17879/freeneuropathology-2023-4790 Kwan, G.T., Frable, B.W., Thompson, A.R., Tresguerres, M., 2022. Optimizing immunostaining of archival fish samples to enhance museum collection potential. Acta Histochem. 124, 151952. https://doi.org/10.1016/j.acthis.2022.151952 Lai, H.M., Liu, A.K.L., Ng, H.H.M., Goldfinger, M.H., Chau, T.W., DeFelice, J., Tilley, B.S., Wong, W.M., Wu, W., Gentleman, S.M., 2018. Next generation histology methods for three-dimensional imaging of fresh and archival human brain tissues. Nat. Commun. 9, 1066. https://doi.org/10.1038/s41467-018-03359-w Lai, H.M., Liu, A.K.L., Ng, W.-L., DeFelice, J., Lee, W.S., Li, H., Li, W., Ng, H.M., Chang, R.C.-C., Lin, B., Wu, W., Gentleman, S.M., 2016. Rationalisation and Validation of an Acrylamide-Free Procedure in Three-Dimensional Histological Imaging. PloS One 11, e0158628. https://doi.org/10.1371/journal.pone.0158628 Larsen, N.Y., Vihrs, N., Møller, J., Sporring, J., Tan, X., Li, X., Ji, G., Rajkowska, G., Sun, F., Nyengaard, J.R., 2022. Layer III pyramidal cells in the prefrontal cortex reveal morphological changes in subjects with depression, schizophrenia, and suicide. Transl. Psychiatry 12, 363. https://doi.org/10.1038/s41398-022-02128-0 Le Beux, Y., Hetenyi, G., Phillips, M.J., 1969. Mitochondrial myelin-like figures: a non-specific reactive process of mitochondrial phospholipid membranes to several stimuli. Z. Zellforsch. Mikrosk. Anat. Vienna Austria 1948 99, 491–506. https://doi.org/10.1007/BF00340941 Leboeuf, C., Ratajczak, P., Zhao, W.-L., François Plassa, L., Court, M., Pisonero, H., Murata, H., Cayuela, J.-M., Ameisen, J.-C., Garin, J., Janin, A., 2008. Long-Term Preservation at Room Temperature of Freeze-Dried Human Tumor Samples Dedicated to Nucleic Acids Analyses. Cell Preserv. Technol. 6, 191–198. https://doi.org/10.1089/cpt.2008.0003 Lembo, T., Niezgoda, M., Velasco-Villa, A., Cleaveland, S., Ernest, E., Rupprecht, C.E., 2006. Evaluation of a direct, rapid immunohistochemical test for rabies diagnosis. Emerg. Infect. Dis. 12, 310–313. https://doi.org/10.3201/eid1202.050812 Lin, Y.-H., Wang, L.-W., Chen, Y.-H., Chan, Y.-C., Hu, S.-H., Wu, S.-Y., Chiang, C.-S., Huang, G.-J., Yang, S.-D., Chu, S.-W., Wang, K.-C., Lin, C.-H., Huang, P.-H., Cheng, H.-J., Chen, B.-C., Chu, L.-A., 2023. Large tissue archiving solution for multiplexed labeling and super-resolution imaging. https://doi.org/10.1101/2022.11.28.518189 Lindberg, M., 1984. Variation in epidermal structure as function of different fixation times: an electron microscopic study. J. Cutan. Pathol. 11, 71–73. https://doi.org/10.1111/j.1600-0560.1984.tb00352.x Liu, A.K.L., Hurry, M.E.D., Ng, O.T.W., DeFelice, J., Lai, H.M., Pearce, R.K.B., Wong, G.T.-C., Chang, R.C.-C., Gentleman, S.M., 2016. Bringing CLARITY to the human brain: visualization of Lewy pathology in three dimensions. Neuropathol. Appl. Neurobiol. 42, 573–587. https://doi.org/10.1111/nan.12293 Liu, J.Y.W., Martinian, L., Thom, M., Sisodiya, S.M., 2010. Immunolabeling recovery in archival, post-mortem, human brain tissue using modified antigen retrieval and the catalyzed signal amplification system. J. Neurosci. Methods 190, 49–56. https://doi.org/10.1016/j.jneumeth.2010.04.020 Liu, X.-B., Schumann, C.M., 2014. Optimization of electron microscopy for human brains with long-term fixation and fixed-frozen sections. Acta Neuropathol. Commun. 2, 42. https://doi.org/10.1186/2051-5960-2-42 Lou, J.J., Mirsadraei, L., Sanchez, D.E., Wilson, R.W., Shabihkhani, M., Lucey, G.M., Wei, B., Singer, E.J., Mareninov, S., Yong, W.H., 2014. A review of room temperature storage of biospecimen tissue and nucleic acids for anatomic pathology laboratories and biorepositories. Clin. Biochem. 47, 267–273. https://doi.org/10.1016/j.clinbiochem.2013.12.011 Luisa, B.G., 1982. The Biology of Crustacea: Volume 1: Systematics, The Fossil Record, And Biogeography. Academic Press. Lundström, Y., Lundström, P., Popova, S.N., Lindblom, R.P.F., Alafuzoff, I., 2019. Detection of Changes in Immunohistochemical Stains Caused by Postmortem Delay and Fixation Time. Appl. Immunohistochem. Mol. Morphol. AIMM 27, 238–245. https://doi.org/10.1097/PAI.0000000000000658 Luyendijk-Elshout, A.M., 1970. Death enlightened. A study of Frederik Ruysch. JAMA 212, 121–126. https://doi.org/10.1001/jama.212.1.121 Lyck, L., Dalmau, I., Chemnitz, J., Finsen, B., Schrøder, H.D., 2008. Immunohistochemical markers for quantitative studies of neurons and glia in human neocortex. J. Histochem. Cytochem. 56, 201–221. https://doi.org/10.1369/jhc.7A7187.2007 Lyon, H., van Deurs, B., Høyer, P.E., Prentø, P., Møller, M., 1991. Tissue Processing: IV. Applied Fixation, in: Lyon, Hans (Ed.), Theory and Strategy in Histochemistry: A Guide to the Selection and Understanding of Techniques. Springer, Berlin, Heidelberg, pp. 187–195. https://doi.org/10.1007/978-3-642-73742-8_13 MacLeod, I.D., Moore, G.I., Gomez, S., 2018. Glycerol impregnation of a formaldehyde fixed Bump-head Sunfish Mola alexandrini. Rec. West. Aust. Mus. 33, 171–180. https://doi.org/10.18195/issn.0312-3162.33(2).2018.171-180 Magoon, E.H., Robb, R.M., 1981. Development of myelin in human optic nerve and tract. A light and electron microscopic study. Arch. Ophthalmol. Chic. Ill 1960 99, 655–659. https://doi.org/10.1001/archopht.1981.03930010655011 Mahesh, S., Tang, K.-C., Raj, M., 2018. Amide Bond Activation of Biological Molecules. Mol. Basel Switz. 23, E2615. https://doi.org/10.3390/molecules23102615 Maki, K.C., Miller, J.W., McCabe, G.P., Raman, G., Kris-Etherton, P.M., 2021. Perspective: Laboratory Considerations and Clinical Data Management for Human Nutrition Randomized Controlled Trials: Guidance for Ensuring Quality and Integrity. Adv. Nutr. Bethesda Md 12, 46–58. https://doi.org/10.1093/advances/nmaa088 Malkovskiy, A.V., Tom, A., Joubert, L.-M., Bao, Z., 2022. Visualization of the distribution of covalently cross-linked hydrogels in CLARITY brain-polymer hybrids for different monomer concentrations. Sci. Rep. 12, 13549. https://doi.org/10.1038/s41598-022-17687-x Maneta-Peyret, L., Compére, P., Moreau, P., Goffinet, G., Cassagne, C., 1999. Immunocytochemistry of Lipids: Chemical Fixatives have Dramatic Effects on the Preservation of Tissue Lipids. Histochem. J. 31, 541–547. https://doi.org/10.1023/A:1003844107756 Marquina, D., Buczek, M., Ronquist, F., Łukasik, P., 2021. The effect of ethanol concentration on the morphological and molecular preservation of insects for biodiversity studies. PeerJ 9, e10799. https://doi.org/10.7717/peerj.10799 Marte, F., Solazzo, C., von Endt, D., Erhardt, D., Tumosa, C.S., 2003. The stability of natural history specimens in fluid-preserved collections, in: 6th International Congress, Cultural Heritage: Context and Conservation in Havana, Cuba. pp. 14–18. Matsuda, Y., Nagao, M., Takatori, T., Niijima, H., Nakajima, M., Iwase, H., Kobayashi, M., Iwadate, K., 1998. Detection of the sarin hydrolysis product in formalin-fixed brain tissues of victims of the Tokyo subway terrorist attack. Toxicol. Appl. Pharmacol. 150, 310–320. https://doi.org/10.1006/taap.1998.8428 McFadden, W.C., Walsh, H., Richter, F., Soudant, C., Bryce, C.H., Hof, P.R., Fowkes, M., Crary, J.F., McKenzie, A.T., 2019. Perfusion fixation in brain banking: a systematic review. Acta Neuropathol. Commun. 7, 146. https://doi.org/10.1186/s40478-019-0799-y McGarvey, K.A., Lee, J.M., Boughner, D.R., 1984. Mechanical suitability of glycerol-preserved human dura mater for construction of prosthetic cardiac valves. Biomaterials 5, 109–117. https://doi.org/10.1016/0142-9612(84)90011-5 McKenzie, A.T., Woodoff-Leith, E., Dangoor, D., Cervera, A., Ressler, H., Whitney, K., Dams-O’Connor, K., Wu, Z., Hillman, E.M.C., Seifert, A.C., Crary, J.F., 2022. Ex situ perfusion fixation for brain banking: a technical report. Free Neuropathol. 3, 22–22. https://doi.org/10.17879/freeneuropathology-2022-4368 Micheva, K.D., Gong, B., Collman, F., Weinberg, R.J., Smith, S.J., Trimmer, J.S., Murray, K.D., 2023. Developing a Toolbox of Antibodies Validated for Array Tomography-Based Imaging of Brain Synapses. eNeuro ENEURO.0290-23.2023. https://doi.org/10.1523/ENEURO.0290-23.2023 Miller, K.L., Stagg, C.J., Douaud, G., Jbabdi, S., Smith, S.M., Behrens, T.E.J., Jenkinson, M., Chance, S.A., Esiri, M.M., Voets, N.L., Jenkinson, N., Aziz, T.Z., Turner, M.R., Johansen-Berg, H., McNab, J.A., 2011. Diffusion imaging of whole, post-mortem human brains on a clinical MRI scanner. NeuroImage 57, 167–181. https://doi.org/10.1016/j.neuroimage.2011.03.070 Mollenhauer, H.H., 1993. Artifacts caused by dehydration and epoxy embedding in transmission electron microscopy. Microsc. Res. Tech. 26, 496–512. https://doi.org/10.1002/jemt.1070260604 Monteiro, S.I.G., 2008. Use of Immunohistochemical Techniques to Investigate Museum Specimens of Neurological Disease. Universidade de Aveiro, Aveiro, Portugal. Morán, M.A., Gómez-Ramos, P., 1992. Cholinesterase histochemistry in the human brain: effect of various fixation and storage conditions. J. Neurosci. Methods 43, 49–54. https://doi.org/10.1016/0165-0270(92)90066-m Morgello, S., Gelman, B.B., Kozlowski, P.B., Vinters, H.V., Masliah, E., Cornford, M., Cavert, W., Marra, C., Grant, I., Singer, E.J., 2001. The National NeuroAIDS Tissue Consortium: a new paradigm in brain banking with an emphasis on infectious disease. Neuropathol. Appl. Neurobiol. 27, 326–335. https://doi.org/10.1046/j.0305-1846.2001.00334.x Mostegl, M.M., Richter, B., Dinhopl, N., Weissenböck, H., 2011. Influence of prolonged formalin fixation of tissue samples on the sensitivity of chromogenic in situ hybridization. J. Vet. Diagn. Invest. 23, 1212–1216. https://doi.org/10.1177/1040638711425584 Murray, E., Cho, J.H., Goodwin, D., Ku, T., Swaney, J., Kim, S.-Y., Choi, H., Park, Y.-G., Park, J.-Y., Hubbert, A., McCue, M., Vassallo, S., Bakh, N., Frosch, M.P., Wedeen, V.J., Seung, H.S., Chung, K., 2015. Simple, Scalable Proteomic Imaging for High-Dimensional Profiling of Intact Systems. Cell 163, 1500–1514. https://doi.org/10.1016/j.cell.2015.11.025 Myers, D.B., Pollock, M., Calder, C.S., 1977. The analysis of collagen in glutaraldehyde-fixed, osmicated peripheral nerve. Acta Neuropathol. (Berl.) 37, 7–12. https://doi.org/10.1007/BF00684533 Nei, T., 1981. Roles of fixatives and cryoprotectants in cryotechniques for electron microscopy: effects of glutaraldehyde and glycerol on structures and functions of platelets. J. Electron Microsc. (Tokyo) 30, 69–71. https://doi.org/10.1093/oxfordjournals.jmicro.a050283 Newman, G.R., Hobot, J.A., 1999. Resins for combined light and electron microscopy: a half century of development. Histochem. J. 31, 495–505. https://doi.org/10.1023/a:1003850921869 Nilsen, T.W., 2014. Preparation of cross-linked cellular extracts with formaldehyde. Cold Spring Harb Protoc. 9, 1001-3. https://doi.org/10.1101/pdb.prot080879 Norris, C.B., Joseph, P.R., Mackiewicz, M.R., Reed, S.M., 2010. Minimizing Formaldehyde Use in the Synthesis of Gold−Silver Core−Shell Nanoparticles. Chem. Mater. 22, 3637–3645. https://doi.org/10.1021/cm9035693 Nowacka, A., Douezan, S., Wadsö, L., Topgaard, D., Sparr, E., 2012. Small polar molecules like glycerol and urea can preserve the fluidity of lipid bilayers under dry conditions. Soft Matter 8, 1482–1491. https://doi.org/10.1039/C1SM06273E Nuovo, G.J., Richart, R.M., 1989. Buffered formalin is the superior fixative for the detection of HPV DNA by in situ hybridization analysis. Am. J. Pathol. 134, 837–842. Nürnberg, E., Vitacolonna, M., Klicks, J., von Molitor, E., Cesetti, T., Keller, F., Bruch, R., Ertongur-Fauth, T., Riedel, K., Scholz, P., Lau, T., Schneider, R., Meier, J., Hafner, M., Rudolf, R., 2020. Routine Optical Clearing of 3D-Cell Cultures: Simplicity Forward. Front. Mol. Biosci. 7, 20. https://doi.org/10.3389/fmolb.2020.00020 Orlowski, P., McConnell, F.K., Payne, S., 2014. A mathematical model of cellular metabolism during ischemic stroke and hypothermia. IEEE Trans. Biomed. Eng. 61, 484–490. https://doi.org/10.1109/TBME.2013.2282603 Otubo, A., Maejima, S., Oti, T., Satoh, K., Ueda, Y., Morris, J.F., Sakamoto, T., Sakamoto, H., 2021. Immunoelectron Microscopic Characterization of Vasopressin-Producing Neurons in the Hypothalamo-Pituitary Axis of Non-Human Primates by Use of Formaldehyde-Fixed Tissues Stored at -25 °C for Several Years. Int. J. Mol. Sci. 22, 9180. https://doi.org/10.3390/ijms22179180 Öztürk, N.C., Koç, T., 2022. Testing the suitability of neuroanatomical tracing method in human fetuses with long years of postmortem delay. Surg. Radiol. Anat. SRA 44, 769–783. https://doi.org/10.1007/s00276-022-02942-7 Paljärvi, L., Garcia, J.H., Kalimo, H., 1979. The efficiency of aldehyde fixation for electron microscopy: stabilization of rat brain tissue to withstand osmotic stress. Histochem. J. 11, 267–276. https://doi.org/10.1007/BF01005026 Papageorgopoulou, C., Rentsch, K., Raghavan, M., Hofmann, M.I., Colacicco, G., Gallien, V., Bianucci, R., Rühli, F., 2010. Preservation of cell structures in a medieval infant brain: a paleohistological, paleogenetic, radiological and physico-chemical study. NeuroImage 50, 893–901. https://doi.org/10.1016/j.neuroimage.2010.01.029 Patel, K.B., Liang, W., Casper, M.J., Voleti, V., Li, W., Yagielski, A.J., Zhao, H.T., Perez Campos, C., Lee, G.S., Liu, J.M., Philipone, E., Yoon, A.J., Olive, K.P., Coley, S.M., Hillman, E.M.C., 2022. High-speed light-sheet microscopy for the in-situ acquisition of volumetric histological images of living tissue. Nat. Biomed. Eng. 6, 569–583. https://doi.org/10.1038/s41551-022-00849-7 Patrono, L.V., Vrancken, B., Budt, M., Düx, A., Lequime, S., Boral, S., Gilbert, M.T.P., Gogarten, J.F., Hoffmann, L., Horst, D., Merkel, K., Morens, D., Prepoint, B., Schlotterbeck, J., Schuenemann, V.J., Suchard, M.A., Taubenberger, J.K., Tenkhoff, L., Urban, C., Widulin, N., Winter, E., Worobey, M., Schnalke, T., Wolff, T., Lemey, P., Calvignac-Spencer, S., 2022. Archival influenza virus genomes from Europe reveal genomic variability during the 1918 pandemic. Nat. Commun. 13, 2314. https://doi.org/10.1038/s41467-022-29614-9 Phillips, J., Laude, A., Lightowlers, R., Morris, C.M., Turnbull, D.M., Lax, N.Z., 2016. Development of passive CLARITY and immunofluorescent labelling of multiple proteins in human cerebellum: understanding mechanisms of neurodegeneration in mitochondrial disease. Sci. Rep. 6, 26013. https://doi.org/10.1038/srep26013 Pikkarainen, M., Martikainen, P., Alafuzoff, I., 2010. The effect of prolonged fixation time on immunohistochemical staining of common neurodegenerative disease markers. J. Neuropathol. Exp. Neurol. 69, 40–52. https://doi.org/10.1097/NEN.0b013e3181c6c13d Pinheiro, T., Mayor, I., Edwards, S., Joven, A., Kantzer, C.G., Kirkham, M., Simon, A., 2021. CUBIC-f: An optimized clearing method for cell tracing and evaluation of neurite density in the salamander brain. J. Neurosci. Methods 348, 109002. https://doi.org/10.1016/j.jneumeth.2020.109002 Ploeger, S., Guldemond, J.M., Feirabend, H.K., Marani, E., 1993. Acidification of human brains stored in fixatives. Eur. J. Morphol. 31, 286–290. Pocivavsek, L., Gavrilov, K., Cao, K.D., Chi, E.Y., Li, D., Lin, B., Meron, M., Majewski, J., Lee, K.Y.C., 2011. Glycerol-induced membrane stiffening: the role of viscous fluid adlayers. Biophys. J. 101, 118–127. https://doi.org/10.1016/j.bpj.2011.05.036 Potaliya, P., Ghatak, S., 2016. Augmenting the Visual Display of Museum Specimen Using Fabrica Acrylic Colors and Its Better Restoration. Indian J. Anat. 5, 269. https://doi.org/10.21088/ija.2320.0022.5316.12 Puchtler, H., Meloan, S.N., 1985. On the chemistry of formaldehyde fixation and its effects on immunohistochemical reactions. Histochemistry 82, 201–204. https://doi.org/10.1007/BF00501395 Purea, A., Webb, A.G., 2006. Reversible and irreversible effects of chemical fixation on the NMR properties of single cells. Magn. Reson. Med. 56, 927–931. https://doi.org/10.1002/mrm.21018 Qiu, W., Zhang, H., Bao, A., Zhu, K., Huang, Y., Yan, X., Zhang, J., Zhong, C., Shen, Y., Zhou, J., Zheng, X., Zhang, L., Shu, Y., Tang, B., Zhang, Z., Wang, G., Zhou, R., Sun, B., Gong, C., Duan, S., Ma, C., 2019. Standardized Operational Protocol for Human Brain Banking in China. Neurosci. Bull. 35, 270–276. https://doi.org/10.1007/s12264-018-0306-7 Radzicka, A., Wolfenden, R., 1996. Rates of Uncatalyzed Peptide Bond Hydrolysis in Neutral Solution and the Transition State Affinities of Proteases. J. Am. Chem. Soc. 118, 6105–6109. https://doi.org/10.1021/ja954077c Rahimi, F., Shepherd, C.E., Halliday, G.M., Geczy, C.L., Raftery, M.J., 2006. Antigen-epitope retrieval to facilitate proteomic analysis of formalin-fixed archival brain tissue. Anal. Chem. 78, 7216–7221. https://doi.org/10.1021/ac060294s Ramos-Vara, J.A., Miller, M.A., 2014. When tissue antigens and antibodies get along: revisiting the technical aspects of immunohistochemistry--the red, brown, and blue technique. Vet. Pathol. 51, 42–87. https://doi.org/10.1177/0300985813505879 Ranly, D.M., 1984. Glutaraldehyde purity and stability: implications for preparation, storage, and use as a pulpotomy agent. Pediatr. Dent. 6, 83–87. Rariy, R.V., Klibanov, A.M., 1997. Correct protein folding in glycerol. Proc. Natl. Acad. Sci. U. S. A. 94, 13520–13523. https://doi.org/10.1073/pnas.94.25.13520 Rauch, J., Muntau, B., Eggert, P., Tappe, D., 2018. Rickettsia typhi as Cause of Fatal Encephalitic Typhus in Hospitalized Patients, Hamburg, Germany, 1940–1944. Emerg. Infect. Dis. 24, 1982–1987. https://doi.org/10.3201/eid2411.171373 Raxworthy, C.J., Smith, B.T., 2021. Mining museums for historical DNA: advances and challenges in museomics. Trends Ecol. Evol. 36, 1049–1060. https://doi.org/10.1016/j.tree.2021.07.009 Rhodes, F.H., Mason, C.W., Sutton, W.R., 1927. Crystallization of Paraffin Wax1. Ind. Eng. Chem. 19, 935–938. https://doi.org/10.1021/ie50212a028 Richtering, W., Saunders, B.R., 2014. Gel architectures and their complexity. Soft Matter 10, 3695–3702. https://doi.org/10.1039/C4SM00208C Richters, C.D., Hoekstra, M.J., van Baare, J., du Pont, J.S., Kamperdijk, E.W., 1996. Morphology of glycerol-preserved human cadaver skin. Burns J. Int. Soc. Burn Inj. 22, 113–116. https://doi.org/10.1016/0305-4179(95)00103-4 Rivera-Quiroz, F.A., Miller, J.A., 2021. Old Brains in Alcohol: The Usability of Legacy Collection Material to Study the Spider Neuroarchitecture. Diversity 13, 601. https://doi.org/10.3390/d13110601 Robards, A.W., Wilson, A.J., 1993. Chapter 5, Basic Biological Preparation Techniques for TEM, in: Procedures in Electron Microscopy. Wiley, England. Romijn, H.J., van Uum, J.F., Breedijk, I., Emmering, J., Radu, I., Pool, C.W., 1999. Double immunolabeling of neuropeptides in the human hypothalamus as analyzed by confocal laser scanning fluorescence microscopy. J. Histochem. Cytochem. Off. J. Histochem. Soc. 47, 229–236. https://doi.org/10.1177/002215549904700211 Rosoklija, G., Mancevski, B., Ilievski, B., Perera, T., Lisanby, S.H., Coplan, J.D., Duma, A., Serafimova, T., Dwork, A.J., 2003. Optimization of Golgi methods for impregnation of brain tissue from humans and monkeys. J. Neurosci. Methods 131, 1–7. https://doi.org/10.1016/j.jneumeth.2003.06.001 Rumjanek, F.D., 2018. Osmolyte Induced Tumorigenesis and Metastasis: Interactions With Intrinsically Disordered Proteins. Front. Oncol. 8, 353. https://doi.org/10.3389/fonc.2018.00353 Sacchini, S., Herráez, P., Arbelo, M., Espinosa de Los Monteros, A., Sierra, E., Rivero, M., Bombardi, C., Fernández, A., 2022. Methodology and Neuromarkers for Cetaceans’ Brains. Vet. Sci. 9, 38. https://doi.org/10.3390/vetsci9020038 Savioz, A., Blouin, J.L., Guidi, S., Antonarakis, S.E., Bouras, C., 1997. A method for the extraction of genomic DNA from human brain tissue fixed and stored in formalin for many years. Acta Neuropathol. (Berl.) 93, 408–413. https://doi.org/10.1007/s004010050632 Schmitz, N., Homberg, F., Berje, J., Burger, J., Hasse, H., 2015. Chemical Equilibrium of the Synthesis of Poly(oxymethylene) Dimethyl Ethers from Formaldehyde and Methanol in Aqueous Solutions. Ind. Eng. Chem. Res. 54, 6409–6417. https://doi.org/10.1021/acs.iecr.5b01148 Schweizer, R., Helms, G., Frahm, J., 2014. Revisiting a historic human brain with magnetic resonance imaging - the first description of a divided central sulcus. Front. Neuroanat. 8, 35. https://doi.org/10.3389/fnana.2014.00035 Sele, M., 2020. Iron deposits in the human post-mortem brain: An electron microscopic study. Graz Medical University, Graz, Austria. Shapson-Coe, A., Januszewski, M., Berger, D.R., Pope, A., Wu, Y., Blakely, T., Schalek, R.L., Li, P.H., Wang, S., Maitin-Shepard, J., Karlupia, N., Dorkenwald, S., Sjostedt, E., Leavitt, L., Lee, D., Bailey, L., Fitzmaurice, A., Kar, R., Field, B., Wu, H., Wagner-Carena, J., Aley, D., Lau, J., Lin, Z., Wei, D., Pfister, H., Peleg, A., Jain, V., Lichtman, J.W., 2021. A connectomic study of a petascale fragment of human cerebral cortex. https://doi.org/10.1101/2021.05.29.446289 Sheaffer, S., Rosoklija, G., Dwork, A.J., 1999. Myelin staining of archival brain tissue. Clin. Neuropathol. 18, 313–317. Shiurba, R.A., Spooner, E.T., Ishiguro, K., Takahashi, M., Yoshida, R., Wheelock, T.R., Imahori, K., Cataldo, A.M., Nixon, R.A., 1998. Immunocytochemistry of formalin-fixed human brain tissues: microwave irradiation of free-floating sections. Brain Res. Brain Res. Protoc. 2, 109–19. https://doi.org/10.1016/s1385-299x(97)00029-9 Sillevis Smitt, P.A., van der Loos, C., Vianney de Jong, J.M., Troost, D., 1993. Tissue fixation methods alter the immunohistochemical demonstrability of neurofilament proteins, synaptophysin, and glial fibrillary acidic protein in human cerebellum. Acta Histochem. 95, 13–21. https://doi.org/10.1016/s0065-1281(11)80381-8 Silvestre, M.A., Yániz, J.L., Peña, F.J., Santolaria, P., Castelló-Ruiz, M., 2021. Role of Antioxidants in Cooled Liquid Storage of Mammal Spermatozoa. Antioxid. Basel Switz. 10, 1096. https://doi.org/10.3390/antiox10071096 Simmons, J., 2019. Storage in Fluid Preservatives, in: Preventive Conservation: Collection Storage. Society for the Preservation of Natural History Collections, New York, NY, pp. 491–507. Sloane, J.A., Hollander, W., Rosene, D.L., Moss, M.B., Kemper, T., Abraham, C.R., 2000. Astrocytic hypertrophy and altered GFAP degradation with age in subcortical white matter of the rhesus monkey. Brain Res. 862, 1–10. https://doi.org/10.1016/s0006-8993(00)02059-x Smarrito-Menozzi, C., Matthey-Doret, W., Devaud-Goumoens, S., Viton, F., 2013. Glycerol, an underestimated flavor precursor in the Maillard reaction. J. Agric. Food Chem. 61, 10225–10230. https://doi.org/10.1021/jf3050044 Snyder, M.L., Lichstein, H.C., 1940. Sodium Azide As An Inhibiting Substance for Gram-Negative Bacteria. J. Infect. Dis. 67, 113–115. https://doi.org/10.1093/infdis/67.2.113 Stoddart, R.W., 1989. Fixatives and preservatives, their effects on tissue, in: Conservation of Natural History Specimens. Spirit Collections. pp. 1–25. Straube, N., Lyra, M.L., Paijmans, J.L.A., Preick, M., Basler, N., Penner, J., Rödel, M.-O., Westbury, M.V., Haddad, C.F.B., Barlow, A., Hofreiter, M., 2021. Successful application of ancient DNA extraction and library construction protocols to museum wet collection specimens. Mol. Ecol. Resour. 21, 2299–2315. https://doi.org/10.1111/1755-0998.13433 Strnad, Š., Strnadová, V., Sýkora, D., Cvačka, J., Maletínská, L., Vrkoslav, V., 2022. MALDI Mass Spectrometry Imaging of Lipids on Free-Floating Brain Sections and Immunohistochemically Colocalized Markers of Neurodegeneration. Methods Mol. Biol. Clifton NJ 2437, 229–239. https://doi.org/10.1007/978-1-0716-2030-4_16 Stumm, M.M., Walker, M.R., Stork, C., Hanoteau, N., Wagner, U., O’Reilly, T.M., 2012. Validation of a postfixation tissue storage and transport medium to preserve histopathology and molecular pathology analyses (total and phosphoactivated proteins, and FISH). Am. J. Clin. Pathol. 137, 429–436. https://doi.org/10.1309/AJCPDZ4FAQ9BUEXC Sufieva, D.A., Fedorova, E.A., Yakovlev, V.S., Kirik, O.V., Tsyba, D.L., Grigorev, I.P., Korzhevskii, D.E., 2023. 25-Year Storage of Human Choroid Plexus in Methyl Salicylate Preserves Its Antigen Immunoreactivity. Turk Patoloji Derg. 39, 109–116. https://doi.org/10.5146/tjpath.2022.01581 Sugiura, K., Koike, S., Suzuki, T., Ogasawara, Y., 2020. Oxidative Formation of Methylglyoxal in Glycerol Preparations during Storage. Biol. Pharm. Bull. 43, 879–883. https://doi.org/10.1248/bpb.b19-01120 Susaki, E.A., Shimizu, C., Kuno, A., Tainaka, K., Li, X., Nishi, K., Morishima, K., Ono, H., Ode, K.L., Saeki, Y., Miyamichi, K., Isa, K., Yokoyama, C., Kitaura, H., Ikemura, M., Ushiku, T., Shimizu, Y., Saito, T., Saido, T.C., Fukayama, M., Onoe, H., Touhara, K., Isa, T., Kakita, A., Shibayama, M., Ueda, H.R., 2020. Versatile whole-organ/body staining and imaging based on electrolyte-gel properties of biological tissues. Nat. Commun. 11, 1982. https://doi.org/10.1038/s41467-020-15906-5 Tam, H., Zhang, W., Infante, D., Parchment, N., Sacks, M., Vyavahare, N., 2017. Fixation of Bovine Pericardium-Based Tissue Biomaterial with Irreversible Chemistry Improves Biochemical and Biomechanical Properties. J Cardiovasc. Transl. Res. 10, 194–205. https://doi.org/10.1007/s12265-017-9733-5 Taylor, D.M., Brown, J.M., Fernie, K., McConnell, I., 1997. The effect of formic acid on BSE and scrapie infectivity in fixed and unfixed brain-tissue. Vet. Microbiol. 58, 167–174. https://doi.org/10.1016/s0378-1135(97)00165-x Tayri-Wilk, T., Slavin, M., Zamel, J., Blass, A., Cohen, S., Motzik, A., Sun, X., Shalev, D.E., Ram, O., Kalisman, N., 2020. Mass spectrometry reveals the chemistry of formaldehyde cross-linking in structured proteins. Nat. Commun. 11, 3128. https://doi.org/10.1038/s41467-020-16935-w Tendler, B.C., Qi, F., Foxley, S., Pallebage-Gamarallage, M., Menke, R.A.L., Ansorge, O., Hurley, S.A., Miller, K.L., 2021. A method to remove the influence of fixative concentration on postmortem T2 maps using a kinetic tensor model. Hum. Brain Mapp. 42, 5956–5972. https://doi.org/10.1002/hbm.25661 Thacker, J.S., Andersen, D., Liang, S., Zieniewicz, N., Trivino-Paredes, J.S., Nahirney, P.C., Christie, B.R., 2021. Unlocking the brain: A new method for Western blot protein detection from fixed brain tissue. J. Neurosci. Methods 348, 108995. https://doi.org/10.1016/j.jneumeth.2020.108995 Thomas, S., Sadanandan, J., Blackburn, S.L., McBride, D.W., Dienel, A., Hong, S., Zeineddine, H.A., Thankamani, P.K., 2022. Glyoxal Fixation Is Optimal for Immunostaining of Brain Vessels, Pericytes and Blood-Brain Barrier Proteins. Int. J. Mol. Sci. 23, 7776. https://doi.org/10.3390/ijms23147776 Thomson, A., 1880. Method of Preserving the Brain. Brain 3, 257–259. https://doi.org/10.1093/brain/3.2.257 Trappe, V., Prasad, V., Cipelletti, L., Segre, P.N., Weitz, D.A., 2001. Jamming phase diagram for attractive particles. Nature 411, 772–775. https://doi.org/10.1038/35081021 Tsutsumi, Y., 2018. Electron Microscopic Study Using Formalin-fixed, Paraffin-embedded Material, with Special Reference to Observation of Microbial Organisms and Endocrine Granules. Acta Histochem. Cytochem. 51, 63–71. https://doi.org/10.1267/ahc.18012 Ueda, H.R., Ertürk, A., Chung, K., Gradinaru, V., Chédotal, A., Tomancak, P., Keller, P.J., 2020. Tissue clearing and its applications in neuroscience. Nat. Rev. Neurosci. 21, 61–79. https://doi.org/10.1038/s41583-019-0250-1 Van Dam, J.A., 2020. Finding A Cure for Hazardous Collections: The Road to Zero Formaldehyde and Ethanol. Collect. Forum 34, 118–123. https://doi.org/10.14351/0831-4985-34.1.118 van der Lem, T., de Bakker, M., Keuck, G., Richardson, M.K., 2021. Wilhelm His Sr. and the development of paraffin embedding. Pathol. 42, 55–61. https://doi.org/10.1007/s00292-021-00947-4 van Duijn, S., Nabuurs, R.J.A., van Rooden, S., Maat-Schieman, M.L.C., van Duinen, S.G., van Buchem, M.A., van der Weerd, L., Natté, R., 2011. MRI artifacts in human brain tissue after prolonged formalin storage. Magn. Reson. Med. 65, 1750–1758. https://doi.org/10.1002/mrm.22758 Van Gieson, I., 1889. Laboratory notes of technical methods for the nervous system. NY Med J 50, 57–60. Van Herp, F., Coenen, T., Geurts, H.P.M., Janssen, G.J.A., Martens, G.J.M., 2005. A fast method to study the secretory activity of neuroendocrine cells at the ultrastructural level. J. Microsc. 218, 79–83. https://doi.org/10.1111/j.1365-2818.2005.01456.x Vásquez-Rivera, A., Oldenhof, H., Dipresa, D., Goecke, T., Kouvaka, A., Will, F., Haverich, A., Korossis, S., Hilfiker, A., Wolkers, W.F., 2018. Use of sucrose to diminish pore formation in freeze-dried heart valves. Sci. Rep. 8, 12982. https://doi.org/10.1038/s41598-018-31388-4 Verhaert, P.D., Walgraeve, H.R., Downer, R.G., 1990. Alkaline phosphatase activity in the brain of the American cockroach Periplaneta americana L. Histochem. J. 22, 628–635. https://doi.org/10.1007/BF01072945 Vitošević, K., Todorović, D., Slović, Ž., Varljen, T., Radaković, I., Radojević, D., Čanović, V., Todorović, M., 2023. The quality of DNA isolated from autopsy formalin-fixed and formalin-fixed paraffin-embedded tissues: study of 1662 samples. Mol. Biol. Rep. 50, 6323–6336. https://doi.org/10.1007/s11033-023-08491-5 Vonsattel, J.P.G., Amaya, M. del P., Cortes, E.P., Mancevska, K., Keller, C.E., 2008. 21st century brain banking practical prerequisites and lessons from the past: the experience of New York Brain Bank – Taub Institute - Columbia University. Cell Tissue Bank. 9, 247–258. https://doi.org/10.1007/s10561-008-9079-y Voogd, J., Feirabend, H.K.P., 1981. Classic methods in neuroanatomy. Methods Neurobiol. 301–364. Vos, D.R.N., Bowman, A.P., Heeren, R.M.A., Balluff, B., Ellis, S.R., 2019. Class-specific depletion of lipid ion signals in tissues upon formalin fixation. Int. J. Mass Spectrom. 446, 116212. https://doi.org/10.1016/j.ijms.2019.116212 Walker, J.F., 1944. Formaldehyde. New York: Reinhold Pub. Wang, N.S., Minassian, H., 1987. The formaldehyde-fixed and paraffin-embedded tissues for diagnostic transmission electron microscopy: a retrospective and prospective study. Hum. Pathol. 18, 715–727. https://doi.org/10.1016/s0046-8177(87)80243-5 Watson, R.E.J., Wiegand, S.J., Clough, R.W., Hoffman, G.E., 1986. Use of cryoprotectant to maintain long-term peptide immunoreactivity and tissue morphology. Peptides 7, 155–159. https://doi.org/10.1016/0196-9781(86)90076-8 Weil, A., 1929. The influence of formalin fixation on the lipoids of the central nervous system. J. Biol. Chem. 83, 601–609. https://doi.org/10.1016/S0021-9258(18)77090-7 Westh, P., 2003. Unilamellar DMPC vesicles in aqueous glycerol: preferential interactions and thermochemistry. Biophys. J. 84, 341–349. https://doi.org/10.1016/S0006-3495(03)74854-1 Wiggermann, V., Endmayr, V., Hernández-Torres, E., Höftberger, R., Kasprian, G., Hametner, S., Rauscher, A., 2023. Quantitative magnetic resonance imaging reflects different levels of histologically determined myelin densities in multiple sclerosis, including remyelination in inactive multiple sclerosis lesions. Brain Pathol. e13150. https://doi.org/10.1111/bpa.13150 Wilcock, G.K., Matthews, S.M., Moss, T., 1990. Comparison of three silver stains for demonstrating neurofibrillary tangles and neuritic plaques in brain tissue stored for long periods. Acta Neuropathol. (Berl.) 79, 566–568. https://doi.org/10.1007/BF00296118 Williamson, D.I., Russell, G., 1965. Ethylene glycol as a preservative for marine organisms. Nature 206, 1370–1371. https://doi.org/10.1038/2061370a0 Woelfle, S., Deshpande, D., Feldengut, S., Roselli, F., Deisseroth, K., Michaelis, J., Boeckers, T.M., Schön, M., 2022. CLARITY increases sensitivity and specificity of fluorescence immunostaining in long-term archived human brain tissue. https://doi.org/10.1101/2022.04.27.489700 Wolff, K.D., Walter, G., Zimmer, C., 1993. Nerve reconstruction with glycerol-preserved allogenic grafts in the rat. Microsurgery 14, 315–322. https://doi.org/10.1002/micr.1920140506 Wong, G., Greenhalgh, T., Westhorp, G., Buckingham, J., Pawson, R., 2013. RAMESES publication standards: realist syntheses. BMC Med. 11, 21. https://doi.org/10.1186/1741-7015-11-21 Wu, K.H.C., Penfold, P.L., Billson, F.A., 2002. Effects of post-mortem delay and storage duration on the expression of GFAP in normal human adult retinae. Clin. Experiment. Ophthalmol. 30, 200–207. https://doi.org/10.1046/j.1442-9071.2002.00515.x Wu, X., Deng, C., Su, Y., Zhang, C., Chen, M., Tian, K., Wu, H., Xu, S., 2022. The effect of prolonged formalin fixation on the expression of proteins in human brain tissues. Acta Histochem. 124, 151879. https://doi.org/10.1016/j.acthis.2022.151879 Xie, R., Chung, J.-Y., Ylaya, K., Williams, R.L., Guerrero, N., Nakatsuka, N., Badie, C., Hewitt, S.M., 2011. Factors influencing the degradation of archival formalin-fixed paraffin-embedded tissue sections. J. Histochem. Cytochem. 59, 356–365. https://doi.org/10.1369/0022155411398488 Zamenhof, S., Grauel, L., Van Marthens, E., Stillinger, R.A., 1972. Quantitative determination of DNA in preserved brains and brain sections. J. Neurochem. 19, 61–68. https://doi.org/10.1111/j.1471-4159.1972.tb01253.x Zhanmu, O., Yang, X., Gong, H., Li, X., 2020. Paraffin-embedding for large volume bio-tissue. Sci. Rep. 10, 12639. https://doi.org/10.1038/s41598-020-68876-5 Zhao, S., Todorov, M.I., Cai, R., -Maskari, R.A., Steinke, H., Kemter, E., Mai, H., Rong, Z., Warmer, M., Stanic, K., Schoppe, O., Paetzold, J.C., Gesierich, B., Wong, M.N., Huber, T.B., Duering, M., Bruns, O.T., Menze, B., Lipfert, J., Puelles, V.G., Wolf, E., Bechmann, I., Ertürk, A., 2020. Cellular and Molecular Probing of Intact Human Organs. Cell 180, 796-812.e19. https://doi.org/10.1016/j.cell.2020.01.030 Zheng, S., Doucette, A.A., 2016. Preventing N- and O-formylation of proteins when incubated in concentrated formic acid. Proteomics 16, 1059–1068. https://doi.org/10.1002/pmic.201500366 Zocher, H., Machado, R.D., 1959. Observations on the solidification of paraffin. Acta Crystallogr. 12, 122–125. https://doi.org/10.1107/S0365110X59000378
Copyright: © 2024 The author(s). This is an open access article distributed under the terms of the Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited, a link to the Creative Commons license is provided, and any changes are indicated. The Creative Commons Public Domain Dedication waiver (https://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated. |