|
|
|
Free Neuropathology 5:5 (2024) |
|
Review |
|
HIV and COVID-19: two pandemics with significant (but different) central nervous system complications |
|
Shino Magaki1, Ting Zhang1, Karam Han1a, Hilda Mirbaha1, William H. Yong2, Cristian Achim3, Gregory Fishbein4, Michael C. Fishbein4, Omai Garner4, Noriko Salamon5, Christopher K. Williams1, Miguel A. Valdes-Sueiras6, Jeffrey J. Hsu7, Theodoros Kelesidis8b, Glenn E. Mathisen9, Helen Lavretsky10, Elyse J. Singer6, Harry V. Vinters1,6,11 |
a Current affiliation: Department of Pathology and Laboratory Medicine, School of Medicine and Public Health, University of Wisconsin-Madison, Madison, WI, USA |
|
Corresponding author: |
|
Submitted: 05 February 2024 |
|
Keywords: HIV, COVID-19, HIV-associated neurocognitive disorders, Long COVID, Post-COVID conditions, Neuropathology |
|
Abstract Human immunodeficiency virus (HIV) and severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) cause significant neurologic disease. Central nervous system (CNS) involvement of HIV has been extensively studied, with well-documented invasion of HIV into the brain in the initial stage of infection, while the acute effects of SARS-CoV-2 in the brain are unclear. Neuropathologic features of active HIV infection in the brain are well characterized whereas neuropathologic findings in acute COVID-19 are largely non-specific. On the other hand, neuropathologic substrates of chronic dysfunction in both infections, as HIV-associated neurocognitive disorders (HAND) and post-COVID conditions (PCC)/long COVID are unknown. Thus far, neuropathologic studies on patients with HAND in the era of combined antiretroviral therapy have been inconclusive, and autopsy studies on patients diagnosed with PCC have yet to be published. Further longitudinal, multidisciplinary studies on patients with HAND and PCC and neuropathologic studies in comparison to controls are warranted to help elucidate the mechanisms of CNS dysfunction in both conditions. |
|
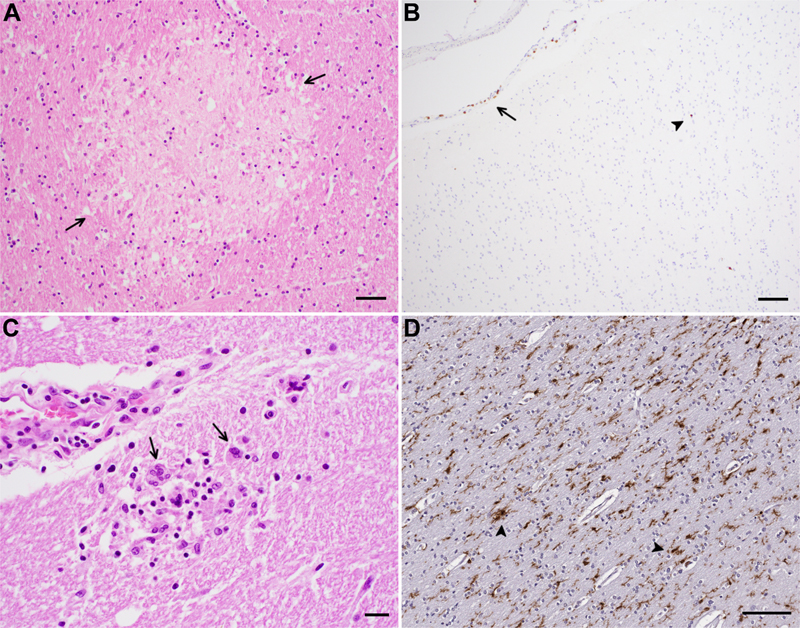
Introduction Human immunodeficiency virus (HIV) and severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) are both human viruses that cause neurologic disease. HIV is a retrovirus and is clearly neurotropic, with tropism for microglial cells in the central nervous system (CNS), infection of which generates downstream effects that injure brain parenchyma. Its clinical and pathologic manifestations are fairly well characterized. By contrast, coronavirus disease 2019 (COVID-19) caused by the novel coronavirus SARS-CoV-2 leads to neuropsychiatric morbidity in a significant percentage of infected patients, but the mechanisms for this are heterogeneous and may include indirect effects on the brain through activation of inflammatory cascades, possible microangiopathic changes and disruption of the neurovascular unit. However, knowledge of HIV neuropathogenesis may be instructive for understanding many aspects of COVID-19 neuropathogenesis. The Brain Research Institute at UCLA sponsored two half-day "Neurovirology Affinity Group" meetings in the spring of 2022 and 2023 at which a multidisciplinary team of representatives from neuropathology, cardiovascular pathology, microbiology, radiology, infectious disease, cardiology, neurology, and psychiatry convened to address the CNS complications of COVID-19, considering what we have learned from the HIV pandemic over the past several decades. The meeting aims were to identify gaps in current knowledge on CNS disease in COVID-19 and HIV and potential directions for future study. This selective review summarizes the main topics discussed and conclusions reached. Human immunodeficiency virus (HIV) An immunodeficiency disease, later named acquired immunodeficiency syndrome (AIDS), started to become recognized in the summer of 1981 in the United States [1,2]. This discovery was followed soon after by the identification of human immunodeficiency virus-1 (HIV-1) as the causative agent [3–5]. At the time, nearly all patients infected with HIV progressed to end stage AIDS with fatal opportunistic infections and malignancies, many involving the CNS [6]. A few years later, HIV-2, primarily restricted to West Africa, was discovered [7–9]. With the advent of combined antiretroviral therapy (cART) in the mid 1990s, HIV has become a chronic disease with near normal life expectancies, at least in industrialized countries [6,10]. There have been approximately 40 million deaths since the beginning of the pandemic and currently 38 million infected worldwide according to the Joint United Nations Programme on HIV/AIDS (UNAIDS) [11]. HIV-1 and HIV-2 are lentiviruses of the retrovirus family and predominantly infect monocytes/macrophages, as with all lentiviruses, but also CD4 T-cells by binding to CD4 and the chemokine co-receptors CXCR4 and CCR5 [12,13]. Other co-receptors such as CCR2 and CCR3 have also been reported to mediate infection in vitro [12,13]. Infection of macrophages (and microglia) is primarily through CCR5 [14,15]. The entrance of virus into the cell may result in productive infection or latent infection [7,12,16,17]. HIV is neuroinvasive, characteristic of lentiviruses, and enters the CNS at the initial stage of infection [13,18–20]. Neurologic manifestations HIV causes neurologic complications, due to opportunistic infections and HIV infection itself, in over half of patients not receiving cART [21,22]. With the advent of cART, the rates of opportunistic infections of the CNS such as by cytomegalovirus, toxoplasma, cryptococcus and progressive multifocal leukoencephalopathy caused by JC virus, as well as primary CNS lymphoma, have decreased in some series [21,23]. HIV-associated neurocognitive disorders (HAND), with revised consensus nomenclature and criteria in 2007 [24], previously referred to as AIDS dementia complex and HIV-1 associated cognitive/motor complex [25–28], comprise a spectrum of cognitive dysfunction associated with HIV infection [24,25]. HAND includes, with increasing severity, asymptomatic neurocognitive impairment (ANI), mild neurocognitive disorder (MND) and HIV-associated dementia (HAD), and continues to affect approximately 33-50 % of treated HIV-positive individuals [24,29–31]. HAD is the most severe and was the most common form of HAND in the pre-cART era but has decreased with the advent of cART, now comprising less than 10 % of those with HAND [6]. However, the overall proportion of individuals with HAND has not changed in the cART era due to a relative increase in the milder forms of HAND, the pathogenesis and neuropathologic substrates of which are unclear [6,31]. The clinical features of HAND have also changed before and after cART [32]. In the pre-cART era, patients commonly presented with extrapyramidal signs (bradykinesia, rigidity and tremor) and more frequent subcortical features with motor dysfunction and speed of processing deficits [26,32–35]. In the cART era, extrapyramidal signs are less common with more cortical features with deficits in learning, memory and executive functioning [32–34,36]. Risk factors for HAND include older age [37], cerebrovascular disease risk factors [38,39], duration of HIV infection and history of AIDS defining illness [40], lower CD4 nadir [40,41], and in treatment naïve patients, burden of HIV DNA in monocyte-enriched peripheral blood cells [42]. Currently, the only treatment for HAND is cART, but cART is effective in only a subset of patients with HAND [32,43]. After the advent of cART, the median survival of HAD patients increased to 38 months compared to 5 months pre-cART [44], and most individuals with HAND on cART remained stable [29,32]. However, even in patients on cART, HAND is associated with shorter survival and is an important cause of morbidity and mortality in the aging HIV-positive population [45–47]. A recent study showed that cognitive decline in HIV positive individuals was not associated with age or markers of HIV disease but rather comorbidities such as hypertension and diabetes [48]. Before cART, brain magnetic resonance imaging (MRI) studies showed accelerated global white matter atrophy and cerebrospinal fluid (CSF) volume increase (corresponding to tissue loss) as well as atrophy of the caudate in HIV-positive individuals [49,50]. Postmortem MRI studies demonstrated that increase in volume of abnormal white matter and decrease in volume of deep gray matter correlated with increasing viral burden as assessed by immunohistochemistry for HIV protein [51]. Even after cART, compared to HIV-negative individuals, HIV-positive individuals show higher rates of white matter atrophy, which correlate with lower CD4 counts [52]. MRI changes can be seen in all degrees of HAND, with individuals with HAND showing greater abnormality in cerebral white matter compared to neurocognitively intact HIV-positive individuals [53]. Utilizing dynamic contrast enhanced perfusion (DCE P) MRI, a marker of capillary permeability, to assess for blood-brain barrier (BBB) disruption, Chaganti et al. showed impaired BBB in the frontal white matter and basal ganglia in virally suppressed patients with HAND compared to controls [54]. Neuropathologic findings Neuropathologic studies have made important contributions to understanding HIV infection in the CNS [7,12,55–57]. HIV has been isolated from the CSF, brain including frontal and temporal lobes, caudate, and cerebellum, spinal cord, and sural nerve from AIDS patients with meningitis, encephalopathy, myelopathy, and peripheral neuropathy [58-60]. Even in asymptomatic patients, HIV specific antibodies can be detected and HIV isolated from the CSF [19,61]. HIV-1 has also been isolated from the brain 15 days after a patient accidentally inoculated himself with HIV-1 infected white blood cells [62]. HIV is thought to enter the CNS predominantly through a Trojan horse-like mechanism in which HIV-1 infected monocytes cross the blood-brain barrier (BBB) and release viral particles in the brain parenchyma [16,19,63] although entry may also be facilitated through a disrupted BBB [64]. HIV-1 has been shown to productively infect brain microglia/macrophages [65–69]. Although HIV-1 DNA and proteins have been detected in other CNS cell types, such as astrocytes and endothelial cells in postmortem brain [17,18,70,71] and shown to infect other cell types in vitro, their role in propagation of CNS infection is unclear [12]. Neuropathologic changes can be seen early in HIV infection [20,72]. In asymptomatic HIV-positive patients who died from other causes, lymphocytic leptomeningitis, perivascular mononuclear cell infiltrates, microgliosis and astrocytic gliosis, which is more prominent in white matter compared to gray matter, myelin pallor, and elevated cytokines, have been seen along with HIV-1 proviral DNA (but no HIV-1 protein) in a subset [20,72,73]. Before the advent of cART, most HIV-positive individuals progressed to AIDS [6], and the majority 70-90 % demonstrated neuropathologic abnormalities at autopsy [27,55,56,68,74–77]. In addition to frequent opportunistic infections and lymphomas seen in the context of immunodeficiency, there are characteristic neuropathologic findings attributed to HIV infection itself [19,27,68,74]. These were summarized in consensus guidelines in 1991 [28] and include HIV encephalitis (HIVE), HIV leukoencephalopathy, lymphocytic (aseptic) leptomeningitis, and diffuse microgliosis and astrogliosis in cerebral gray matter ("diffuse poliodystrophy") [7,56,78]. HIV encephalitis, seen in 10-63 % of autopsies, is characterized by microglial nodules and multinucleated giant cells (Figure 1) and/or evidence of HIV in the brain [57,79,80] typically affecting the white matter most severely, followed by the deep gray matter and then cortex [27,28,65,80]. Multinucleated giant cells, often seen in a perivascular distribution and more commonly in the subcortical white matter or deep gray matter, are the hallmark finding in HIVE and result from virus-induced fusion of macrophages which have been shown to contain HIV protein [65,81–83].
Figure 1. Neuropathologic findings in COVID-19 and HIV infection. A) Acute/subacute microinfarct (arrows) in the internal capsule and B) sparse leptomeningeal (arrow) and parenchymal (arrowhead) T lymphocytes highlighted by CD3 immunohistochemistry in patients with COVID-19. C) HIV encephalitis with multinucleated giant cells (arrows) and D) microglial activation (arrowheads) highlighted by immunohistochemistry for Iba-1 in patients with HIV infection. Scale bars: A = 50 μm, B,D = 100 μm, C = 20 μm In the post cART era, the majority of HIV positive individuals show no HIV related neuropathology at autopsy, although HIVE has been reported in 8-25 % of case series with equal or even increased frequency post cART in some series [84–87]. Opportunistic infections, albeit decreased in frequency after cART, are still seen [84–92]. HIVE indicates active viral replication and is an important pathologic substrate of HAD [93], but not all subjects with cognitive impairment have HIVE and not all patients with HIVE have cognitive impairment, which was seen before but especially after the advent of cART [56,68,81,84,88,94]. In the post cART era, HIV associated neuropathologic findings have not correlated with cognitive impairment [84]. AIDS patients demonstrate cortical atrophy and neuronal apoptosis [95,96], but neither neuronal apoptosis [97] nor neuronal loss in the frontal and temporal cortices of patients with AIDS [98] correlate with dementia. On the other hand, decreased cortical synaptic density and dendritic complexity were demonstrated even with mild cognitive impairment and have been shown to correlate with severity of cognitive impairment [99–101]. Disruption of the BBB has been seen in patients both with AIDS with and without dementia as well as with and without HIVE [64,102]. Several gene expression profiling studies performed on the brains of HIV positive individuals have found upregulation of endothelial cell type transcripts [103] and dysregulation of microglial transcripts [104] in patients with HAND but without HIVE and increased expression of a subset of cytokines in white matter [103–107]. Although HIV antigen can be detected throughout the CNS including the cerebral hemispheres, cerebellum, brainstem, and spinal cord using various methods such as immunohistochemistry, in situ hybridization (ISH), PCR, and electron microscopy, the highest levels of HIV proteins, RNA and DNA, primarily in macrophages, microglia and multinucleated giant cells, have been seen in deep gray matter and hippocampus [67,81,108,109]. Even with viral suppression on cART, low levels of HIV-1 RNA and anti-HIV antibodies can be detected in the CSF [110,111]. HIV-1 DNA, including intact proviruses, and RNA have been found in the brains of virally suppressed HIV-positive individuals in microglia/macrophages [112–114]. These reservoirs have been postulated to cause chronic inflammation and neuronal dysfunction [114,115], and microglial activation and increased cytokine levels in the brain have been shown to correlate with HAND [94,116–119]. However, although levels of HIV-1 DNA are on average higher in brains from patients with AIDS compared to patients without AIDS, there is overlap [18,117]. Brain viral load correlates poorly with neuropathologic changes or cognitive impairment [94,117]. Furthermore, Gelman et al. have shown that subjects with HAND and HIVE have higher brain HIV RNA and DNA levels than patients without HAND, but individuals with HAND without HIVE show no difference compared to patients without HAND [88]. Thus, complex mechanisms beyond viral replication in the brain likely underlie HAND, especially in the post cART era (Figure 2) [56,84,120,121].
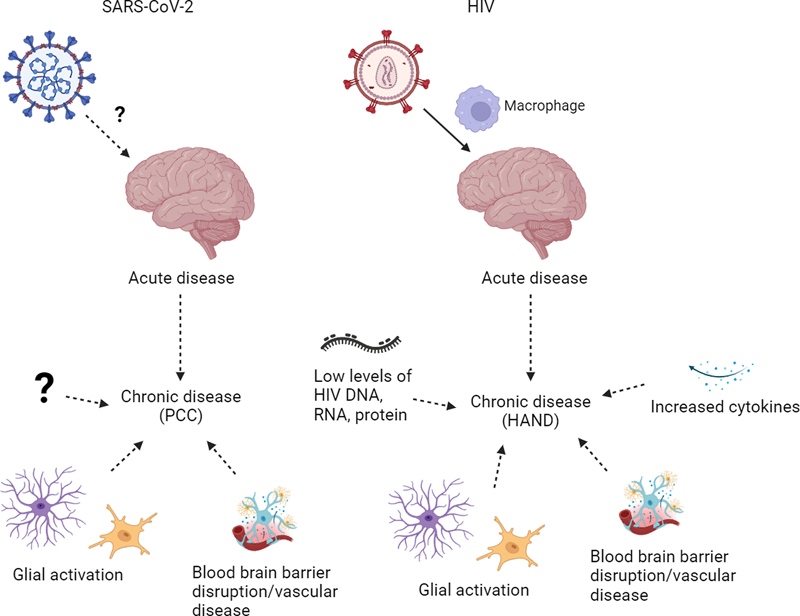
Figure 2. Potential mechanisms of neurologic disease in acute/chronic HIV infection and COVID-19/PCC. HAND, HIV-associated neurocognitive disorders; PCC, post-COVID conditions. Figure was created using BioRender. Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) A few weeks after a cluster of viral pneumonia cases, later called COVID-19, was first described in December of 2019 in China [122–124], the causative agent was determined to be a new human pathogen, the coronavirus later named severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) [123]. Several vaccines were rapidly developed within a year and a significant proportion of the population of many countries, mostly high-income, have been vaccinated, albeit with large differences among countries [122,125,126]. However, there have been approximately 6.9 million deaths and over 750 million infected worldwide according to the World Health Organization (WHO) [127]. Coronaviruses are classified into four distinct genera: Alpha, Beta, Gamma and Deltacoronavirus [128]. SARS-CoV-2 and SARS-CoV belong to the same species, severe acute respiratory syndrome-related coronavirus (SARSr-CoV), which is of the subgenus Sarbecovirus, genus Betacoronavirus which also includes Middle East respiratory syndrome coronavirus (MERS-CoV), of the subfamily Orthocoronaviridae [128,129]. During its spread several SARS-CoV-2 variants have emerged, termed "variants of concern", including Alpha, Gamma, Delta, and Omicron, that have caused disease of similar severity except for Omicron which has been associated with lower rates of hospitalization [128,130]. SARS-CoV-2 predominantly infects nasal or upper respiratory epithelium and sustentacular cells of the olfactory mucosa [131–134] with a decreasing gradient of infection from the proximal to distal respiratory tract [135]. As with SARS-CoV, SARS-CoV-2 uses angiotensin converting enzyme 2 (ACE2) as the obligate receptor for host cell entry through binding of the viral spike (S) protein, proteolytically activated by transmembrane protease/serine subfamily member 2 (TMPRSS2), although ACE2-bound virus can also enter the cell via clathrin-mediated endocytosis without TMPRSS2 [129,136–140]. ACE2 is a human homologue of ACE, and both ACE and ACE2 play important roles in the renin-angiotensin system [141]. ACE2 mRNA has been reported in many different organs including, at least in some studies, the brain [142–145]. However, mRNA and protein levels of ACE2 have been shown to be discordant [146]. ACE2 protein has been detected in several cell types including ciliated cells of the nasal mucosa and bronchus, cardiomyocytes, enterocytes of the gastrointestinal tract, gallbladder epithelium, kidney proximal tubule epithelium, and Sertoli and Leydig cells of the testis [143,146,147]. In the brain, ACE2 protein expression has been seen by immunohistochemistry in vascular smooth muscle cells and/or endothelial cells/pericytes in some studies but not others [143,147–151]. Few studies have reported ACE2 immunopositivity in choroid plexus, ependymal cells, meningothelial cells, and neurons in the medulla [151,152]. Neurologic manifestations Approximately a third of patients with acute COVID-19 exhibit neurologic and psychiatric manifestations, most commonly fatigue, alterations in consciousness, impairment in smell and taste, seizures, myalgia, anxiety, and stroke [153–156]. Ischemic stroke is seen in approximately 2 % of COVID-19 patients [153–156], and occurs in younger individuals compared to ischemic stroke in non-COVID-19 patients [157]. Moreover, patients with COVID-19 have increased risk for ischemic and hemorrhagic stroke compared to patients with influenza or other upper respiratory tract infections [155,158], and increased risk for ischemic stroke and myocardial infarct compared to the background population in Sweden [159]. Neurologic manifestations have been associated with increased mortality, especially in older individuals [154,156]. Neurologic and psychiatric complications are also seen as part of the constellation of post-COVID conditions (PCC), also known as post-acute sequelae of COVID-19 (PASC) or long COVID, defined by the WHO as persistent symptoms usually 3 months from onset of COVID-19 with symptoms lasting at least 2 months with no alternative diagnosis [160–162]. Most frequent neurologic manifestations of PCC include fatigue and cognitive impairment, impaired concentration or "brain fog", which has been shown to correlate with abnormal brain activation on task-activated blood oxygenation level-dependent functional MRI (BOLD-fMRI) [163], as well as psychiatric disorders such as post-traumatic stress disorder, anxiety and depression [164,165]. Similar complications were seen in the SARS 2003 pandemic [166], and post-viral syndromes occur in other viruses [167]. Cognitive decline and psychiatric symptoms have also been described after serious illness requiring admission to the intensive care unit such as sepsis [168,169]. However, examining the electronic health records of the US Department of Veterans Affairs, Al-Aly et al. found a higher burden of both pulmonary and extrapulmonary disease, including neuropsychiatric and cardiovascular disorders, in patients who had been hospitalized for COVID-19 and survived for at least 30 days after admission compared to individuals hospitalized for influenza [170]. A different study using electronic health records of health care organizations mostly in the US found that neurologic and psychiatric diagnoses are more common in the 6 months following the diagnosis of COVID-19 compared to other respiratory infections such as influenza [155]. PCC can occur regardless of the severity of acute disease but is more severe in hospitalized patients, and those with comorbidities appear to be at increased risk not only for acute disease but also for PCC [160,164,171]. Vaccination may be less effective at preventing PCC than severe acute COVID-19 [172]. Further clinical phenotyping of PCC, as with HAND in HIV, may be helpful in further defining research nosology [24]. Brain MRI findings in patients with acute COVID-19 presenting with neurologic symptoms include acute/subacute infarct, which is the most common finding, abnormalities of the olfactory bulb, white matter abnormalities, cerebral microbleeds, gray matter abnormalities, leptomeningeal enhancement, acute disseminated encephalomyelitis (ADEM) and ADEM-like lesions, intracerebral hemorrhage, and posterior reversible encephalopathy syndrome (PRES) [173,174]. In 18F-FDG PET studies, subacute COVID-19 patients have shown hypometabolism in the frontoparietal cortex, which correlated with cognitive performance, with reduction in hypometabolism along with improvement in cognitive performance approximately 6 months after onset of symptoms [175,176]. In a large longitudinal study examining participants in the UK Biobank who had undergone repeat imaging, a decrease in gray matter thickness in the orbitofrontal cortex and parahippocampal gyrus as well as decrease in global brain size were seen between two scans in those who had been infected by SARS-CoV-2, likely with different variants and averaging 4-5 months after infection, compared to those who had not been infected by SARS-CoV-2 [177]. Greater cognitive decline was also seen in the SARS-CoV-2 positive group [177]. Subsequently, Du et al. found that male patients demonstrated reduced gray matter thickness in the parietal and occipital cortices and hippocampal volume after Omicron infection compared to before infection [178]. In patients diagnosed with PCC, gray matter volume loss has been associated with cognitive dysfunction [179]. Neuropathologic findings Brain autopsy examination of patients who died from COVID-19 have shown a variety of neuropathologic findings, although many are non-specific [180–184]. Most commonly seen are acute hypoxic-ischemic changes with neuronal eosinophilia, acute/subacute infarcts (Figure 1) with macrophage infiltration and neovascularization, hemorrhage [149,185–191], microthrombi [192–194], astrogliosis and microglial activation occasionally with microglial nodules and neuronophagia especially in the cerebellum and brainstem [185,195–199], and T-lymphocytic infiltration, predominantly sparse in a perivascular distribution [180,182,185,186,193, 195,200,201]. Similar features including hypoxic-ischemic injury, microinfarcts, hemorrhage, and sparse lymphocytic infiltrate as well as fibrinogen leakage were seen between Delta, Omicron and non-Delta/non-Omicron variants [202]. Other findings described in patients who died from acute COVID-19 infection include perivascular hemosiderin deposition/leakage [182,193], though perivascular hemosiderin deposition is a common finding in autopsy brains, especially in older individuals, and can be seen in both brains with and without significant vascular disease [182,203–206]. They have also been shown to correspond to a subset of cerebral microbleeds on MRI [207]. Megakaryocytes in cortical capillaries and in an infarct were also reported in acute COVID-19, but capillary megakaryocytes can been seen in the setting of lung injury due to a variety of causes [208–210]. Uncommon findings include multifocal necrotizing leukoencephalopathy [185], acute encephalitis with lymphohistiocytic infiltrate and hemorrhage [200], and ADEM/acute hemorrhagic leukoencephalopathy (AHLE) and ADEM/AHLE-like pathology [149,195,211–213]. Some comorbid neuropathologic findings described include other infections such as HSV-1 encephalitis [185] and bacterial infection [193], neurodegenerative diseases including Alzheimer disease and Lewy body disease [185,195], and cerebrovascular disease such as cerebral amyloid angiopathy [210], not surprising given the older patient age in some series [185,195,200,214]. Few studies have compared brains from COVID-19 patients to controls [152,197,198,200]. No differences in frequencies of hypoxic-ischemic changes, infarcts or hemorrhages were seen between COVID-19 and non-COVID-19 patients in one study [200]. In another study, perivascular and leptomeningeal T-lymphocytic infiltrates were seen in both COVID-19 patients and in controls with sepsis /systemic inflammation, and microglial, activation in the pons in COVID-19 patients was greater as compared to controls without sepsis but similar to controls with sepsis [152]. However, Lee et al. showed that the brains of COVID-19 patients demonstrate increased leakage of fibrinogen, consistent with BBB disruption, complement activation and microthrombi compared to controls, supportive of neurovascular injury [198,215]. Leakage of fibrinogen and other serum proteins in the brain in COVID-19 have also been reported [150,202]. Several gene expression profiling studies on various regions of the brain from COVID-19 patients have shown downregulation of neuronal and synaptic pathways in the olfactory bulb and amygdala [200], dysregulation that overlap with aging and neurodegenerative diseases in the frontal cortex [145,216], metabolic dysregulation in the brainstem [198], and inflammation in the choroid plexus [145,217]. Very few autopsy studies have examined patients who recovered from COVID-19 prior to death; these have also shown microglial activation/macrophages [195,197]. One of the studies included two patients with mild COVID-19 symptoms who died 4-5 and 10 weeks after testing positive for COVID-19 from causes unrelated to the infection [197]. In these subjects microglial activation/macrophages were also seen, similar to findings in patients with acute COVID-19 in the brainstem and cerebellum but to a milder degree in the cerebral white matter [197]. So far there have been no autopsy studies of patients diagnosed with PCC (long COVID). In a brief look at other pandemics, hypoxic- ischemic changes, infarcts, hemorrhage, microglial activation, and ADEM-like lesions were also seen in autopsies of patients who died from the novel influenza A H1N1 virus in 2009 [218]. The few autopsies reported on SARS-CoV, the etiologic agent responsible for the SARS pandemic which emerged in 2002 [219], described hypoxic-ischemic changes and gliosis in the brain [220,221]. There are only rare reports of SARS-CoV detected in the CSF [222,223] and in brain using PCR, ISH, immunohistochemistry, and electron microscopy [220–222,224] with isolation of SARS-CoV reported from the brain of one patient [221]. The "brain isolation" may have reflected isolation of virus from brain vasculature. Although several studies have detected SARS-CoV-2 RNA in the brain using PCR or transcriptomic analysis [152,185,193,200,225–227], including in the affected area of a case of hemorrhagic encephalitis possibly attributed to COVID-19 [200], other studies have not detected viral RNA or protein in the brains of patients with COVID-19 using a variety of methods [145,182,186,192,195,197,198,201,216,217], and SARS-CoV-2 has never been cultured from brain [131,180]. Few studies have performed cellular localization of viral RNA using ISH or viral protein using immunohistochemistry and most have shown that they are restricted to vessels/near vessels such as in endothelium, within macrophages or near capillaries [148,182,225,228]. There are rare reports of viral protein or RNA in brain tissue or cranial nerves [148,182,185,229–231], including a report by Thakur et al. which found low levels of viral RNA by RT-PCR but no viral RNA or protein by RNAscope and immunohistochemistry [185]. Viral RNA was seen in the adventitia of a meningeal blood vessel outside the medulla in one case [185]. Because of frequent anosmia/hyposmia and olfactory dysfunction in COVID-19, the suspicion is that the virus may enter the brain via the olfactory route through the cribriform plate, similar to neurotropic viruses [232–234]. Several studies have examined the olfactory bulbs [185,193,201,230,231,235]. Most showed variable degrees of T-lymphocytic infiltration and gliosis without presence of viral protein, but variability in presence of viral RNA in human tissue [185,193,201,230,231,235]. These discrepancies may partly be due to differences in patient cohorts and methods employed including the usage of different commercial antibodies. Some antibodies, including a widely used spike antibody (Abcam 3A2), have subsequently been shown to label vessels and neurons in both COVID-19 and control patients or have high background [145,180,197,236]. Viral nucleocapsid antigen and RNA have only rarely been detected in CSF in acute disease [237–240]. CSF analysis often shows slightly increased white blood cell counts as well as elevated albumin and protein levels and an elevated albumin quotient, which suggests disruption of the BBB [238]. Similar to the brain, SARS-CoV-2 has never been cultured from CSF even when virus could be grown from paired nasopharyngeal swabs [131]. In electron microscopy studies, virions have mainly been identified in the respiratory tract, and reports of ultrastructural detection of virus outside the respiratory tract are controversial due to potential misinterpretation of virus-like particles [236,241–243]. Thus, it is unclear whether SARS-CoV-2 is neurotropic or neuroinvasive, and mechanisms other than direct infection likely contribute to both acute and chronic forms of the disease (Figure 2) [167,232]. Conclusions and next steps HIV and SARS-CoV-2 are associated with significant neurologic morbidity but with differing pathogenesis underlying acute and chronic disease. In acute disease, HIV directly invades the CNS while SARS-CoV-2 may exert its effects more indirectly through systemic inflammation. In chronic disease, HIV persists in the brain despite cART, and sequelae of low-level infection may contribute to HAND [115]. PCC may also be associated with sequelae of inflammation, but in both diseases, complex interactions with comorbidities likely underlie neurologic manifestations [48,160]. Furthermore, both PCC and HAND have significant components of psychiatric disease in which functional abnormalities are thought to predominate over structural abnormalities [244,245]. Mechanisms of aging related neurodegeneration have also been implicated. Brain lysates from COVID-19 patients have shown activation of TGF-β signaling and increased oxidative stress as well as activation of pathways leading to tau hyperphosphorylation associated with Alzheimer disease [246]. One promising avenue of investigation is the cerebral vasculature, as vascular disease and BBB dysfunction have been implicated to play a significant role in HAND and acute COVID-19 [54,64,102,198]. Furthermore, it is well established that vascular disease plays a significant role in aging and neurodegenerative diseases [247], pathways which have been implicated in COVID-19 as well as HAND [145,216,248,249]. The following are potential "next steps" to elucidate the mechanisms underlying the neurologic manifestations in HAND and COVID-19/PCC discussed at the two meetings:
Funding Brain Research Institute, David Geffen School of Medicine, University of California Los Angeles, Los Angeles, CA, USA Disclosure statement None of the authors has any conflict of interest to disclose. References 1. Ruelas DS, Greene WC. An integrated overview of HIV-1 latency. Cell 2013;155:519–29. https://doi.org/10.1016/j.cell.2013.09.044. 2. Gottlieb MS, Schroff R, Schanker HM, Weisman JD, Fan PT, Wolf RA, et al. Pneumocystis carinii pneumonia and mucosal candidiasis in previously healthy homosexual men: evidence of a new acquired cellular immunodeficiency. N Engl J Med 1981;305:1425–31. https://doi.org/10.1056/NEJM198112103052401. 3. Greene WC. A history of AIDS: Looking back to see ahead. Eur J Immunol 2007;37:94–102. https://doi.org/10.1002/eji.200737441. 4. Barré-Sinoussi F, Chermann JC, Rey F, Nugeyre MT, Chamaret S, Gruest J, et al. Isolation of a T-lymphotropic retrovirus from a patient at risk for acquired immune deficiency syndrome (AIDS). Science 1983;220:868–71. https://doi.org/10.1126/science.6189183. 5. Popovic M, Sarngadharan MG, Read E, Gallo RC. Detection, isolation, and continuous production of cytopathic retroviruses (HTLV-III) from patients with AIDS and pre-AIDS. Science 1984;224:497–500. https://doi.org/10.1126/science.6200935. 6. Saylor D, Dickens AM, Sacktor N, Haughey N, Slusher B, Pletnikov M, et al. HIV-associated neurocognitive disorder — pathogenesis and prospects for treatment. Nat Rev Neurol 2016;12:234–48. https://doi.org/10.1038/nrneurol.2016.27. 7. Scaravilli F, Bazille C, Gray F. Neuropathologic contributions to understanding AIDS and the central nervous system. Brain Pathol 2007;17:197–208. https://doi.org/10.1111/j.1750-3639.2007.00047.x. 8. Clavel F, Guétard D, Brun-Vézinet F, Chamaret S, Rey MA, Santos-Ferreira MO, et al. Isolation of a new human retrovirus from West African patients with AIDS. Science 1986;233:343–6. https://doi.org/10.1126/science.2425430. 9. Clavel F, Mansinho K, Chamaret S, Guetard D, Favier V, Nina J, et al. Human immunodeficiency virus type 2 infection associated with AIDS in West Africa. N Engl J Med 1987;316:1180–5. https://doi.org/10.1056/NEJM198705073161903. 10. Trickey A, May MT, Vehreschild J-J, Obel N, Gill MJ, Crane HM, et al. Survival of HIV-positive patients starting antiretroviral therapy between 1996 and 2013: a collaborative analysis of cohort studies. Lancet HIV 2017;4:e349–56. https://doi.org/10.1016/S2352-3018(17)30066-8. 11. Joint United Nations Programme on HIV/AIDS. UNAIDS Data 2022. https://www.unaids.org/sites/default/files/media_asset/data-book-2022_en.pdf. Accessed: June 28, 2023. 12. Bissel SJ, Wiley CA. Human immunodeficiency virus infection of the brain: pitfalls in evaluating infected/affected cell populations. Brain Pathol 2004;14:97–108. https://doi.org/10.1111/j.1750-3639.2004.tb00503.x. 13. Patrick MK, Johnston JB, Power C. Lentiviral neuropathogenesis: comparative neuroinvasion, neurotropism, neurovirulence, and host neurosusceptibility. J Virol 2002;76:7923–31. https://doi.org/10.1128/jvi.76.16.7923-7931.2002. 14. Stevenson M. HIV-1 pathogenesis. Nat Med 2003;9:853–60. https://doi.org/10.1038/nm0703-853. 15. Albright A V., Shieh JTC, Itoh T, Lee B, Pleasure D, O’Connor MJ, et al. Microglia express CCR5, CXCR4, and CCR3, but of these, CCR5 is the principal coreceptor for human immunodeficiency virus type 1 dementia isolates. J Virol 1999;73:205–13. https://doi.org/10.1128/jvi.73.1.205-213.1999. 16. González-Scarano F, Martín-García J. The neuropathogenesis of AIDS. Nat Rev Immunol 2005;5:69–81. https://doi.org/10.1038/nri1527. 17. Saito Y, Sharer LR, Epstein LG, Michaels J, Mintz M, Louder M, et al. Overexpression of nef as a marker for restricted HIV‐1 infection of astrocytes in postmortem pediatric central nervous tissues. Neurology 1994;44:474–474. https://doi.org/10.1212/WNL.44.3_PART_1.474. 18. An SF, Groves M, Giometto B, Beckett AAJ, Scaravilli F. Detection and localisation of HIV-1 DNA and RNA in fixed adult AIDS brain by polymerase chain reaction/in situ hybridisation technique. Acta Neuropathol 1999;98:481–7. https://doi.org/10.1007/s004010051113. 19. Resnick L, Berger JR, Shapshak P, Tourtellotte WW. Early penetration of the blood-brain-barrier by HIV. Neurology 1988;38:9–14. https://doi.org/10.1212/WNL.38.1.9. 20. Gray F, Scaravilli F, Everall I, Chretien F, An S, Boche D, et al. Neuropathology of early HIV-1 infection. Brain Pathol 1996;6:1–15. https://doi.org/10.1111/j.1750-3639.1996.tb00775.x. 21. Valcour V, Sithinamsuwan P, Letendre S, Ances B. Pathogenesis of HIV in the central nervous system. Curr HIV/AIDS Rep 2011;8:54–61. https://doi.org/10.1007/s11904-010-0070-4. 22. Clifford DB. HIV-associated neurocognitive disorder. Curr Opin Infect Dis 2017;30:117–22. https://doi.org/10.1097/QCO.0000000000000328. 23. Tan IL, Smith BR, von Geldern G, Mateen FJ, McArthur JC. HIV-associated opportunistic infections of the CNS. Lancet Neurol 2012;11:605–17. https://doi.org/10.1016/S1474-4422(12)70098-4. 24. Antinori A, Arendt G, Becker JT, Brew BJ, Byrd DA, Cherner M, et al. Updated research nosology for HIV-associated neuro cognitive disorders. Neurology 2007;69:1789–99. https://doi.org/10.1212/01.WNL.0000287431.88658.8b. 25. Janssen RS, Cornblath DR, Epstein LG, Foa RP, McArthur JC, Price RW, et al. Nomenclature and research case definitions for neurologic manifestations of human immunodeficiency virus-type 1 (HIV-1) infection. Report of a Working Group of the American Academy of Neurology AIDS Task Force. Neurology 1991;41:778–85. https://doi.org/10.1212/wnl.41.6.778. 26. Navia BA, Cho BD, Price RW. The AIDS dementia complex: I. Clinical features. Ann Neurol 1986;19:517–24. https://doi.org/10.1002/ana.410190603. 27. Ho DD, Bredesen DE, Vinters HV, Daar ES. The acquired immunodeficiency syndrome (AIDS) dementia complex. Ann Intern Med 1989;111:400–10. https://doi.org/10.7326/0003-4819-111-5-400. 28. Budka H, Wiley CA, Kleihues P, Artigas J, Asbury AK, Cho ES, et al. HIV-associated disease of the nervous system: review of nomenclature and proposal for neuropathology-based terminology. Brain Pathol 1991;1:143–52. https://doi.org/10.1111/j.1750-3639.1991.tb00653.x. 29. Sacktor N, Skolasky RL, Seaberg E, Munro C, Becker JT, Martin E, et al. Prevalence of HIV-associated neurocognitive disorders in the Multicenter AIDS Cohort Study. Neurology 2016;86:334–40. https://doi.org/10.1212/WNL.0000000000002277. 30. Clifford DB, Ances BM. HIV-associated neurocognitive disorder. Lancet Infect Dis 2013;13:976–86. https://doi.org/10.1016/S1473-3099(13)70269-X. 31. Heaton RK, Clifford DB, Franklin DR Jr, Woods SP, Ake C, Vaida F, et al. HIV-associated neurocognitive disorders persist in the era of potent antiretroviral therapy. Neurology 2010;75:2087–96. https://doi.org/10.1212/WNL.0b013e318200d727. 32. Sacktor N. Changing clinical phenotypes of HIV-associated neurocognitive disorders. J Neurovirol 2018;24:141–5. https://doi.org/10.1007/s13365-017-0556-6. 33. Heaton RK, Franklin DR, Ellis RJ, McCutchan JA, Letendre SL, LeBlanc S, et al. HIV-associated neurocognitive disorders before and during the era of combination antiretroviral therapy: Differences in rates, nature, and predictors. J Neurovirol 2011;17:3–16. https://doi.org/10.1007/s13365-010-0006-1. 34. Sacktor N, Robertson K. Evolving clinical phenotypes in HIV-associated neurocognitive disorders. Curr Opin HIV AIDS 2014;9:517–20. https://doi.org/10.1097/COH.0000000000000102. 35. Smail RC, Brew BJ. HIV-associated neurocognitive disorder. Handb Clin Neurol 2018;152:75-97. https://doi.org/10.1016/B978-0-444-63849-6.00007-4. 36. Cysique LA, Maruff P, Brew BJ. Prevalence and pattern of neuropsychological impairment in human immunodeficiency virus-infected/acquired immunodeficiency syndrome (HIV/AIDS) patients across pre- and post-highly active antiretroviral therapy eras: a combined study of two cohorts. J Neurovirol 2004;10:350–7. https://doi.org/10.1080/13550280490521078. 37. Valcour V, Shikuma C, Shiramizu B, Watters M, Poff P, Selnes O, et al. Higher frequency of dementia in older HIV-1 individuals: the Hawaii aging with HIV-1 cohort. Neurology 2004;63:822–7. https://doi.org/10.1212/01.WNL.0000134665.58343.8D. 38. Becker JT, Kingsley L, Mullen J, Cohen B, Martin E, Miller EN, et al. Vascular risk factors, HIV serostatus, and cognitive dysfunction in gay and bisexual men. Neurology 2009;73:1292–9. https://doi.org/10.1212/WNL.0b013e3181bd10e7. 39. McCutchan JA, Marquie-Beck JA, FitzSimons CA, Letendre SL, Ellis RJ, Heaton RK, et al. Role of obesity, metabolic variables, and diabetes in HIV-associated neurocognitive disorder. Neurology 2012;78:485–92. https://doi.org/10.1212/WNL.0b013e3182478d64. 40. Bhaskaran K, Mussini C, Antinori A, Walker AS, Dorrucci M, Sabin C, et al. Changes in the incidence and predictors of human immunodeficiency virus-associated dementia in the era of highly active antiretroviral therapy. Ann Neurol 2008;63:213–21. https://doi.org/10.1002/ana.21225. 41. Ellis RJ, Badiee J, Vaida F, Letendre S, Heaton RK, Clifford D, et al. CD4 nadir is a predictor of HIV neurocognitive impairment in the era of combination antiretroviral therapy. AIDS 2011;25:1747–51. https://doi.org/10.1097/QAD.0b013e32834a40cd. 42. Valcour VG, Ananworanich J, Agsalda M, Sailasuta N, Chalermchai T, Schuetz A, et al. HIV DNA reservoir increases risk for cognitive disorders in cART-naïve patients. PLoS One 2013;8:e70164. https://doi.org/10.1371/journal.pone.0070164. 43. Robertson KR, Robertson WT, Ford S, Watson D, Fiscus S, Harp AG, et al. Highly active antiretroviral therapy improves neurocognitive functioning. J Acquir Immune Defic Syndr 2004;36:562–6. https://doi.org/10.1097/00126334-200405010-00003. 44. Dore GJ, McDonald A, Li Y, Kaldor JM, Brew BJ. Marked improvement in survival following AIDS dementia complex in the era of highly active antiretroviral therapy. AIDS 2003;17:1539–45. https://doi.org/10.1097/00002030-200307040-00015. 45. Sevigny JJ, Albert SM, McDermott MP, Schifitto G, McArthur JC, Sacktor N, et al. An evaluation of neurocognitive status and markers of immune activation as predictors of time to death in advanced HIV infection. Arch Neurol 2007;64:97–102. https://doi.org/10.1001/archneur.64.1.97. 46. Vivithanaporn P, Heo G, Gamble J, Krentz HB, Hoke A, Gill MJ, et al. Neurologic disease burden in treated HIV/AIDS predicts survival: a population-based study. Neurology 2010;75:1150–8. https://doi.org/10.1212/WNL.0b013e3181f4d5bb. 47. Rosenthal J, Tyor W. Aging, comorbidities, and the importance of finding biomarkers for HIV-associated neurocognitive disorders. J Neurovirol 2019;25:673–85. https://doi.org/10.1007/s13365-019-00735-0. 48. Heaton RK, Ellis RJ, Tang B, Marra CM, Rubin LH, Clifford DB, et al. Twelve-year neurocognitive decline in HIV is associated with comorbidities, not age: a CHARTER study. Brain 2023;146:1121–31. https://doi.org/10.1093/brain/awac465. 49. Stout JC, Ellis RJ, Jernigan TL, Archibald SL, Abramson I, Wolfson T, et al. Progressive cerebral volume loss in human immunodeficiency virus infection: a longitudinal volumetric magnetic resonance imaging study. Arch Neurol 1998;55:161–8. https://doi.org/10.1001/archneur.55.2.161. 50. Jernigan TL, Grant I, Hesselink JR, Atkinson JH, Velin RA, McCutchan JA, et al. Magnetic resonance imaging morphometric analysis of cerebral volume loss in human immunodeficiency virus infection. Arch Neurol 1993;50:250–5. https://doi.org/10.1001/archneur.1993.00540030016007. 51. Heindel WC, Jernigan TL, Archibald SL, Achim CL, Masliah E, Wiley CA. The relationship of quantitative brain magnetic resonance imaging measures to neuropathologic indexes of human immunodeficiency virus infection. Arch Neurol 1994;51:1129–35. https://doi.org/10.1001/archneur.1994.00540230067015. 52. Cardenas V, Meyerhoff D, Studholme C, Kornak J, Rothlind J, Lampiris H, et al. Evidence for ongoing brain injury in human immunodeficiency viruspositive patients treated with antiretroviral therapy. J Neurovirol 2009;15:324–33. https://doi.org/10.1080/13550280902973960. 53. Alakkas A, Ellis RJ, Watson CW-M, Umlauf A, Heaton RK, Letendre S, et al. White matter damage, neuroinflammation, and neuronal integrity in HAND. J Neurovirol 2019;25:32–41. https://doi.org/10.1007/s13365-018-0682-9. 54. Chaganti J, Marripudi K, Staub LP, Rae CD, Gates TM, Moffat KJ, et al. Imaging correlates of the blood-brain barrier disruption in HIV-associated neurocognitive disorder and therapeutic implications. AIDS 2019;33:1843–52. https://doi.org/10.1097/QAD.0000000000002300. 55. Bell JE. An update on the neuropathology of HIV in the HAART era. Histopathology 2004;45:549–59. https://doi.org/10.1111/j.1365-2559.2004.02004.x. 56. Morgello S. HIV neuropathology. Handb Clin Neurol 2018;152:3–19. https://doi.org/10.1016/B978-0-444-63849-6.00002-5. 57. Shaw GM, Harper ME, Hahn BH, Epstein LG, Gajdusek DC, Price RW, et al. HTLV-III infection in brains of children and adults with AIDS encephalopathy. Science 1985;227:177–82. https://doi.org/10.1126/science.298142. 58. Ho DD, Sarngadharan MG, Resnick L, Dimarzoveronese F, Rota TR, Hirsch MS. Primary human T-lymphotropic virus type III infection. Ann Intern Med 1985;103:880–3. https://doi.org/10.7326/0003-4819-103-6-880. 59. Levy JA, Shimabukuro J, Hollander H, Mills J, Kaminsky L. Isolation of AIDS-associated retroviruses from cerebrospinal fluid and brain of patients with neurological symptoms. Lancet 1985;2:585–8. 60. Gray F, Chrétien F, Vallat-Decouvelaere AV, Scaravilli F. The changing pattern of HIV neuropathology in the HAART era. J Neuropathol Exp Neurol 2003;62:429–40. https://doi.org/10.1093/jnen/62.5.429. 61. Elovaara I, Seppälä I, Poutiainen E, Suni J, Valle SL. Intrathecal humoral immunologic response in neurologically symptomatic and asymptomatic patients with human immunodeficiency virus infection. Neurology 1988;38:1451–6. https://doi.org/10.1212/WNL.38.9.1451. 62. Davis LE, Hjelle BL, Miller VE, Palmer DL, Llewellyn AL, Merlin TL, et al. Early viral brain invasion in iatrogenic human immunodeficiency virus infection. Neurology 1992;42:1736–9. https://doi.org/10.1212/wnl.42.9.1736. 63. Haase AT. Pathogenesis of lentivirus infections. Nature 1986;322:130–6. https://doi.org/10.1038/322130a0. 64. Petito CK, Cash KS. Blood-brain barrier abnormalities in the acquired immunodeficiency syndrome: immunohistochemical localization of serum proteins in postmortem brain. Ann Neurol 1992;32:658–66. https://doi.org/10.1002/ana.410320509. 65. Wiley CA, Schrier RD, Nelson JA, Lampert PW, Oldstone MB. Cellular localization of human immunodeficiency virus infection within the brains of acquired immune deficiency syndrome patients. Proc Natl Acad Sci U S A 1986;83:7089–93. https://doi.org/10.1073/pnas.83.18.7089. 66. Wiley CA, Achim CL, Christopherson C, Kidane Y, Kwok S, Masliah E, Mellors J, Radhakrishnan L, Wang G, Soontornniyomkij V. HIV mediates a productive infection of the brain. AIDS 1999;13:2055–9. https://doi.org/10.1097/00002030-199910220-00007. 67. Koenig S, Gendelman HE, Orenstein JM, Canto MC, Pezeshkpour H, Yungbluth M, et al. Detection of AIDS virus in macrophages in brain tissue from AIDS patients with encephalopathy. Science 1986;233:1089–98. https://doi.org/10.1126/science.3016903. 68. Navia BA, Cho ES, Petito CK PR. The AIDS dementia complex: II. Neuropathology. Ann Neurol 1986;19:525–35. https://doi.org/10.1002/ana.410190603. 69. Cosenza MA, Zhao ML, Si Q, Lee SC. Human brain parenchymal microglia express CD14 and CD45 and are productively infected by HIV-1 in HIV-1 encephalitis. Brain Pathol 2003;12:442–55. https://doi.org/10.1111/j.1750-3639.2002.tb00461.x. 70. Churchill MJ, Wesselingh SL, Cowley D, Pardo CA, McArthur JC, Brew BJ, et al. Extensive astrocyte infection is prominent in human immunodeficiency virus - associated dementia. Ann Neurol 2009;66:253–8. https://doi.org/10.1002/ana.21697. 71. Thompson KA, Cherry CL, Bell JE, McLean CA. Brain cell reservoirs of latent virus in presymptomatic HIV-infected individuals. Am J Pathol 2011;179:1623–9. https://doi.org/10.1016/j.ajpath.2011.06.039. 72. Gray F, Lescs MC, Keohane C, Paraire F, Marc B, Durigon M, et al. Early brain changes in HIV infection: neuropathological study of 11 HIV seropositive, non-AIDS cases. J Neuropathol Exp Neurol 1992;51:177–85. https://doi.org/10.1097/00005072-199203000-00007. 73. An SF, Ciardi A, Giometto B, Scaravilli T, Gray F, Scaravilli F. Investigation on the expression of major histocompatibility complex class II and cytokines and detection of HIV-1 DNA within brains of asymptomatic and symptomatic HIV-1-positive patients. Acta Neuropathol 1996;91:494–503. https://doi.org/10.1007/s004010050457. 74. Anders KH, Guerra WF, Tomiyasu U, Verity MA, Vinters HV. The neuropathology of AIDS. UCLA experience and review. Am J Pathol 1986;124:537–58. PMC1888341. 75. Vago L, Trabattoni G, Lechi A, Cristina S, Budka H. Neuropathology of AIDS dementia. A review after 205 post mortem examinations. Acta Neurol (Napoli). 1990 Feb;12:32-5. 76. Walsh K, Thompson W, Megyesi J, Wiley CA, Hammond R. HIV-1/AIDS neuropathology in a Canadian teaching centre. Can J Neurol Sci 2004;31:235–41. https://doi.org/10.1017/S0317167100053889. 77. Snider WD, Simpson DM, Nielsen S, Gold JW, Metroka CE, Posner JB. Neurological complications of acquired immune deficiency syndrome: analysis of 50 patients. Ann Neurol 1983;14:403–18. https://doi.org/10.1002/ana.410140404. 78. Budka H, Costanzi G, Cristina S, Lechi A, Parravicini C, Trabattoni R, et al. Brain pathology induced by infection with the human immunodeficiency virus (HIV). A histological, immunocytochemical, and electron microscopical study of 100 autopsy cases. Acta Neuropathol 1987;75:185–98. https://doi.org/10.1007/bf00687080. 79. Budka H. Multinucleated giant cells in brain: a hallmark of the acquired immune deficiency syndrome (AIDS). Acta Neuropathol 1986;69:253–8. https://doi.org/10.1007/BF00688301. 80. Sharer LR, Cho ES, Epstein LG. Multinucleated giant cells and HTLV-III in AIDS encephalopathy. Hum Pathol 1985;16:760. https://doi.org/10.1016/S0046-8177(85)80245-8. 81. Brew BJ, Rosenblum M, Cronin K, Price RW. AIDS dementia complex and HIV‐1 brain infection: clinical‐virological correlations. Ann Neurol 1995;38:563–70. https://doi.org/10.1002/ana.410380404. 82. Shieh JT, Martín J, Baltuch G, Malim MH, González-Scarano F. Determinants of syncytium formation in microglia by human immunodeficiency virus type 1: role of the V1/V2 domains. J Virol 2000;74:693–701. https://doi.org/10.1128/jvi.74.2.693-701.2000. 83. Budka H. Neuropathology of human immunodeficiency virus infection. Brain Pathol 1991;1:163–75. https://doi.org/10.1111/j.1750-3639.1991.tb00656.x. 84. Everall I, Vaida F, Khanlou N, Lazzaretto D, Achim C, Letendre S, et al. Cliniconeuropathologic correlates of human immunodeficiency virus in the era of antiretroviral therapy. J Neurovirol 2009;15:360–70. https://doi.org/10.3109/13550280903131915. 85. Langford TD, Letendre SL, Larrea GJ, Masliah E. Changing patterns in the neuropathogenesis of HIV during the HAART era. Brain Pathol 2003;13:195–210. https://doi.org/10.1111/j.1750-3639.2003.tb00019.x. 86. Masliah E, DeTeresa RM, Mallory ME, Hansen LA. Changes in pathological findings at autopsy in AIDS cases for the last 15 years. AIDS 2000;14:69–74. https://doi.org/10.1097/00002030-200001070-00008. 87. Morgello S, Mahboob R, Yakoushina T, Khan S, Hague K. Autopsy findings in a human immunodeficiency virus-infected population over 2 decades: influences of gender, ethnicity, risk factors, and time. Arch Pathol Lab Med 2002;126:182–90. https://doi.org/10.5858/2002-126-0182-AFIAHI. 88. Gelman BB, Lisinicchia JG, Morgello S, Masliah E, Commins D, Achim CL, et al. Neurovirological correlation with HIV-associated neurocognitive disorders and encephalitis in a HAART-era cohort. J Acquir Immune Defic Syndr 2013;62:487–95. https://doi.org/10.1097/QAI.0b013e31827f1bdb. 89. Jellinger KA, Setinek U, Drlicek M, Böhm G, Steurer A, Lintner F. Neuropathology and general autopsy findings in AIDS during the last 15 years. Acta Neuropathol 2000;100:213–20. https://doi.org/10.1007/s004010000245. 90. Umlauf A, Soontornniyomkij B, Sundermann EE, Gouaux B, Ellis RJ, Levine AJ, et al. Risk of developing cerebral β-amyloid plaques with posttranslational modification among HIV-infected adults. AIDS 2019;33:2157–66. https://doi.org/10.1097/QAD.0000000000002336. 91. Gray F, Keohane C. The neuropathology of HIV infection in the era of highly active antiretroviral therapy (HAART). Brain Pathol 2003;13:79–83. https://doi.org/10.1111/j.1750-3639.2003.tb00008.x. 92. Vago L, Bonetto S, Nebuloni M, Duca P, Carsana L, Zerbi P, et al. Pathological findings in the central nervous system of AIDS patients on assumed antiretroviral therapeutic regimens: retrospective study of 1597 autopsies. AIDS 2002;16:1925–8. https://doi.org/10.1097/00002030-200209270-00009. 93. Wiley CA, Achim C. Human immunodeficiency virus encephalitis is the pathological correlate of dementia in acquired immunodeficiency syndrome. Ann Neurol 1994;36:673–6. https://doi.org/10.1002/ana.410360422. 94. Glass JD, Fedor H, Wesselingh SL, McArthur JC. Immunocytochemical quantitation of human immunodeficiency virus in the brain: correlations with dementia. Ann Neurol 1995;38:755–62. https://doi.org/10.1002/ana.410380510. 95. Gray F, Haug H, Chimelli L, Geny C, Gaston A, Scaravilli F, et al. Prominent cortical atrophy with neuronal loss as correlate of human immunodeficiency virus encephalopathy. Acta Neuropathol 1991;82:229–33. https://doi.org/10.1007/BF00294450. 96. Adle-Biassette H, Levy Y, Colombel M, Poron F, Natchev S, Keohane C, et al. Neuronal apoptosis in HIV infection in adults. Neuropathol Appl Neurobiol 1995;21:218–27. https://doi.org/10.1111/j.1365-2990.1995.tb01053.x. 97. Adle-Biassette H, Chrétien F, Wingertsmann L, Héry C, Ereau T, Scaravilli F, et al. Neuronal apoptosis does not correlate with dementia in HIV infection but is related to microglial activation and axonal damage. Neuropathol Appl Neurobiol 1999;25:123–33. https://doi.org/10.1046/j.1365-2990.1999.00167.x. 98. Everall IP, Glass JD, McArthur J, Spargo E, Lantos P. Neuronal density in the superior frontal and temporal gyri does not correlate with the degree of human immunodeficiency virus-associated dementia. Acta Neuropathol 1994;88:538–44. https://doi.org/10.1007/BF00296490. 99. Everall I, Heaton R, Marcotte T, Ellis R, McCutchan J, Atkinson J, et al. Cortical synaptic density is reduced in mild to moderate human immunodeficiency virus neurocognitive disorder. HIV Neurobehavioral Research Center. Brain Pathol 1999;9:209–17. https://doi.org/10.1111/j.1750-3639.1999.tb00219.x. 100. Masliah E, Heaton RK, Marcotte TD, Ellis RJ, Wiley CA, Mallory M, et al. Dendritic injury is a pathological substrate for human immunodeficiency virus-related cognitive disorders. Ann Neurol 1997;42:963–72. https://doi.org/10.1002/ana.410420618. 101. Masliah E, Achim CL, Ge N, DeTeresa R, Terry RD, Wiley CA. Spectrum of human immunodeficiency virus–associated neocortical damage. Ann Neurol 1992;32:321–9. https://doi.org/10.1002/ana.410320304. 102. Power C, Kong PA, Crawford TO, Wesselingh S, Glass JD, McArthur JC, et al. Cerebral white matter changes in acquired immunodeficiency syndrome dementia: alterations of the blood-brain barrier. Ann Neurol 1993;34:339–50. https://doi.org/10.1002/ana.410340307. 103. Gelman BB, Chen T, Lisinicchia JG, Soukup VM, Carmical JR, Starkey JM, et al. The National NeuroAIDS Tissue Consortium brain gene array: two types of HIV-associated neurocognitive impairment. PLoS One 2012;7:e46178. https://doi.org/10.1371/journal.pone.0046178. 104. Ginsberg SD, Alldred MJ, Gunnam SM, Schiroli C, Lee SH, Morgello S, et al. Expression profiling suggests microglial impairment in human immunodeficiency virus neuropathogenesis. Ann Neurol 2018;83:406–17. https://doi.org/10.1002/ana.25160. 105. Judge M, Parker E, Naniche D, Le Souëf P. Gene expression: the key to understanding HIV-1 infection? Microbiol Mol Biol Rev 2020;84:e00080-19. https://doi.org/10.1128/mmbr.00080-19. 106. Sanna PP, Repunte-Canonigo V, Masliah E, Lefebvre C. Gene expression patterns associated with neurological disease in human HIV infection. PLoS One 2017;12:e0175316. https://doi.org/10.1371/journal.pone.0175316. 107. Solomon IH, De Girolami U, Chettimada S, Misra V, Singer EJ, Gabuzda D. Brain and liver pathology, amyloid deposition, and interferon responses among older HIV-positive patients in the late HAART era. BMC Infect Dis 2017;17:151. https://doi.org/10.1186/s12879-017-2246-7. 108. Achim CL, Wang R, Miners DK, Wiley CA. Brain viral burden in HIV infection. J Neuropathol Exp Neurol 1994;53:284–94. https://doi.org/10.1097/00005072-199405000-00010. 109. Wiley CA, Soontornniyomkij V, Radhakrishnan L, Masliah E, Mellors J, Hermann SA, et al. Distribution of brain HIV load in AIDS. Brain Pathol 1998;8:277–84. https://doi.org/10.1111/j.1750-3639.1998.tb00153.x. 110. Anderson AM, Muñoz-Moreno JA, McClernon DR, Ellis RJ, Cookson D, Clifford DB, et al. Prevalence and correlates of persistent HIV-1 RNA in cerebrospinal fluid during antiretroviral therapy. J Infect Dis 2017;215:105–13. https://doi.org/10.1093/INFDIS/JIW505. 111. Burbelo PD, Price RW, Hagberg L, Hatano H, Spudich S, Deeks SG, et al. Anti-human immunodeficiency virus antibodies in the cerebrospinal fluid: evidence of early treatment impact on central nervous system reservoir? J Infect Dis 2018;217:1024–32. https://doi.org/10.1093/infdis/jix662. 112. Ko A, Kang G, Hattler JB, Galadima HI, Zhang J, Li Q, et al. Macrophages but not astrocytes harbor HIV DNA in the brains of HIV-1-infected aviremic individuals on suppressive antiretroviral therapy. J Neuroimmune Pharmacol 2019;14:110–9. https://doi.org/10.1007/s11481-018-9809-2. 113. Smith DB, Simmonds P, Bell JE. Brain viral burden, neuroinflammation and neurodegeneration in HAART-treated HIV positive injecting drug users. J Neurovirol 2014;20:28–38. https://doi.org/10.1007/s13365-013-0225-3. 114. Cochrane CR, Angelovich TA, Byrnes SJ, Waring E, Guanizo AC, Trollope GS, et al. Intact HIV proviruses persist in the brain despite viral suppression with ART. Ann Neurol 2022;92:532–44. https://doi.org/10.1002/ana.26456. 115. Desplats P, Dumaop W, Smith D, Adame A, Everall I, Letendre S, et al. Molecular and pathologic insights from latent HIV-1 infection in the human brain. Neurology 2013;80:1415–23. https://doi.org/10.1212/WNL.0b013e31828c2e9e. 116. Glass JD, Wesselingh SL, Selnes OA, McArthur JC. Clinical-neuropathologic correlation in HIV-associated dementia. Neurology 1993;43:2230–7. https://doi.org/10.1212/wnl.43.11.2230. 117. An SF, Giometto B, Scaravilli F. HIV-1 DNA in brains in AIDS and pre-AIDS: correlation with the stage of disease. Ann Neurol 1996;40:611–7. https://doi.org/10.1002/ana.410400411. 118. Achim CL, Morey MK, Wiley CA. Expression of major histocompatibility complex and HIV antigens within the brains of AIDS patients. AIDS 1991;5:535–41. https://doi.org/10.1097/00002030-199105000-00009. 119. Tyor WR, Glass JD, Griffin JW, Becker PS, McArthur JC, Bezman L, et al. Cytokine expression in the brain during the acquired immunodeficiency syndrome. Ann Neurol 1992;31:349–60. https://doi.org/10.1002/ana.410310402. 120. Ellis R, Langford D, Masliah E. HIV and antiretroviral therapy in the brain: Neuronal injury and repair. Nat Rev Neurosci 2007;8:33–44. https://doi.org/10.1038/nrn2040. 121. Magaki SD, Vinters HV, Williams CK, Mareninov S, Khanlou N, Said J, et al. Neuropathologic findings in elderly HIV-positive individuals. 2022;81:565-76. https://doi.org/10.1093/jnen/nlac040. 122. Carvalho T, Krammer F, Iwasaki A. The first 12 months of COVID-19: a timeline of immunological insights. Nat Rev Immunol 2021;21:245–56. https://doi.org/10.1038/s41577-021-00522-1. 123. Zhu N, Zhang D, Wang W, Li X, Yang B, Song J, et al. A novel coronavirus from patients with pneumonia in China, 2019. N Engl J Med 2020;382:727–33. https://doi.org/10.1056/nejmoa2001017. 124. Huang C, Wang Y, Li X, Ren L, Zhao J, Hu Y, et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020;395:497–506. https://doi.org/10.1016/S0140-6736(20)30183-5. 125. Mathieu E, Ritchie H, Ortiz-Ospina E, Roser M, Hasell J, Appel C, et al. A global database of COVID-19 vaccinations. Nat Hum Behav 2021;5:947–53. https://doi.org/10.1038/s41562-021-01122-8. 126. Bilinski A, Thompson K, Emanuel E. COVID-19 and excess all-cause mortality in the US and 20 comparison countries, June 2021-March 2022. JAMA 2023;329:92–4. https://doi.org/10.1001/jama.2022.21795. 127. World Health Organization. WHO coronavirus (COVID-19) dashboard. https://covid19.who.int/. Accessed: July 26, 2023. 128. Telenti A, Hodcroft EB, Robertson DL. The evolution and biology of SARS-CoV-2 variants. Cold Spring Harb Perspect Med 2022;12:a041390. https://doi.org/10.1101/cshperspect.a041390. 129. Jackson CB, Farzan M, Chen B, Choe H. Mechanisms of SARS-CoV-2 entry into cells. Nat Rev Mol Cell Biol 2022;23:3–20. https://doi.org/10.1038/s41580-021-00418-x. 130. Esper FP, Adhikari TM, Tu ZJ, Cheng YW, El-Haddad K, Farkas DH, et al. Alpha to Omicron: disease severity and clinical outcomes of major SARS-CoV-2 variants. J Infect Dis 2023;227:344–52. https://doi.org/10.1093/infdis/jiac411 . 131. Khan M, Clijsters M, Choi S, Backaert W, Claerhout M, Couvreur F, et al. Anatomical barriers against SARS-CoV-2 neuroinvasion at vulnerable interfaces visualized in deceased COVID-19 patients. Neuron 2022;110:3919-35. https://doi.org/10.1016/j.neuron.2022.11.007. 132. Khan M, Yoo SJ, Clijsters M, Backaert W, Vanstapel A, Speleman K, et al. Visualizing in deceased COVID-19 patients how SARS-CoV-2 attacks the respiratory and olfactory mucosae but spares the olfactory bulb. Cell 2021;184:5932-49. https://doi.org/10.1016/j.cell.2021.10.027. 133. Lamers MM, Haagmans BL. SARS-CoV-2 pathogenesis. Nat Rev Microbiol 2022;20:270–84. https://doi.org/10.1038/s41579-022-00713-0. 134. Ahn JH, Kim JM, Hong SP, Choi SY, Yang MJ, Ju YS, et al. Nasal ciliated cells are primary targets for SARS-CoV-2 replication in the early stage of COVID-19. J Clin Invest 2021;131:e148517. https://doi.org/10.1172/JCI148517. 135. Hou YJ, Okuda K, Edwards CE, Martinez DR, Asakura T, Dinnon KH, et al. SARS-CoV-2 reverse genetics reveals a variable infection gradient in the respiratory tract. Cell 2020;182:429-46. https://doi.org/10.1016/j.cell.2020.05.042. 136. Li W, Moore MJ, Vasilieva N, Sui J, Wong SK, Berne MA, et al. Angiotensin-converting enzyme 2 is a functional receptor for the SARS coronavirus. Nature 2003;426:450–4. https://doi.org/10.1038/nature02145. 137. Zhou P, Yang XL, Wang XG, Hu B, Zhang L, Zhang W, et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020;579:270–3. https://doi.org/10.1038/s41586-020-2012-7. 138. Millet JK, Jaimes JA, Whittaker GR. Molecular diversity of coronavirus host cell entry receptors. FEMS Microbiol Rev 2021;45:fuaa057. https://doi.org/10.1093/femsre/fuaa057. 139. Hoffmann M, Kleine-Weber H, Schroeder S, Krüger N, Herrler T, Erichsen S, et al. SARS-CoV-2 cell entry depends on ACE2 and TMPRSS2 and is blocked by a clinically proven protease inhibitor. Cell 2020;181:271-80. https://doi.org/10.1016/j.cell.2020.02.052. 140. Letko M, Marzi A, Munster V. Functional assessment of cell entry and receptor usage for SARS-CoV-2 and other lineage B betacoronaviruses. Nat Microbiol 2020;5:562–9. https://doi.org/10.1038/s41564-020-0688-y. 141. Donoghue M, Hsieh F, Baronas E, Godbout K, Gosselin M, Stagliano N, et al. A novel angiotensin-converting enzyme-related carboxypeptidase (ACE2) converts angiotensin I to angiotensin 1-9. Circ Res 2000;87:E1-9. https://doi.org/10.1161/01.res.87.5.e1. 142. Harmer D, Gilbert M, Borman R, Clark KL. Quantitative mRNA expression profiling of ACE 2, a novel homologue of angiotensin converting enzyme. FEBS Lett 2002;532:107–10. https://doi.org/10.1016/S0014-5793(02)03640-2. 143. Hikmet F, Méar L, Edvinsson Å, Micke P, Uhlén M, Lindskog C. The protein expression profile of ACE2 in human tissues. Mol Syst Biol 2020;16:e9610. https://doi.org/10.15252/msb.20209610. 144. Chen R, Wang K, Yu J, Howard D, French L, Chen Z, et al. The spatial and cell-type distribution of SARS-CoV-2 receptor ACE2 in the human and mouse brains. Front Neurol 2021;11:573095. https://doi.org/10.3389/fneur.2020.573095. 145. Yang AC, Kern F, Losada PM, Agam MR, Maat CA, Schmartz GP, et al. Dysregulation of brain and choroid plexus cell types in severe COVID-19. Nature 2021;595:565–71. https://doi.org/10.1038/s41586-021-03710-0. 146. Wang Y, Wang Y, Luo W, Huang L, Xiao J, Li F, et al. A comprehensive investigation of the mRNA and protein level of ACE2, the putative receptor of SARS-CoV-2, in human tissues and blood cells. Int J Med Sci 2020;17:1522–31. https://doi.org/10.7150/ijms.46695. 147. Hamming I, Timens W, Bulthuis MLC, Lely AT, Navis GJ, van Goor H. Tissue distribution of ACE2 protein, the functional receptor for SARS coronavirus. A first step in understanding SARS pathogenesis. J Pathol 2004;203:631–7. https://doi.org/10.1002/path.1570. 148. Nuovo GJ, Magro C, Shaffer T, Awad H, Suster D, Mikhail S, et al. Endothelial cell damage is the central part of COVID-19 and a mouse model induced by injection of the S1 subunit of the spike protein. Ann Diagn Pathol 2021;51:151682. https://doi.org/10.1016/j.anndiagpath.2020.151682. 149. Bryce C, Grimes Z, Pujadas E, Ahuja S, Beasley MB, Albrecht R, et al. Pathophysiology of SARS-CoV-2: the Mount Sinai COVID-19 autopsy experience. Mod Pathol 2021;34:1456–67. https://doi.org/10.1038/s41379-021-00793-y. 150. Bocci M, Oudenaarden C, Sàenz‐sardà X, Simrén J, Edén A, Sjölund J, et al. Infection of brain pericytes underlying neuropathology of COVID‐19 patients. Int J Mol Sci 2021;22:11622. https://doi.org/10.3390/ijms222111622. 151. Lindskog C, Méar L, Virhammar J, Fällmar D, Kumlien E, Hesselager G, et al. Protein expression profile of ACE2 in the normal and COVID-19-affected human brain. J Proteome Res 2022;21:2137–45. https://doi.org/10.1021/acs.jproteome.2c00184. 152. Deigendesch N, Sironi L, Kutza M, Wischnewski S, Fuchs V, Hench J, et al. Correlates of critical illness-related encephalopathy predominate postmortem COVID-19 neuropathology. Acta Neuropathol 2020;140:583–6. https://doi.org/10.1007/s00401-020-02213-y. 153. Mao L, Jin H, Wang M, Hu Y, Chen S, He Q, et al. Neurologic manifestations of hospitalized patients with coronavirus disease 2019 in Wuhan, China. JAMA Neurol 2020;77:683–90. https://doi.org/10.1001/jamaneurol.2020.1127. 154. Misra S, Kolappa K, Prasad M, Radhakrishnan D, Thakur KT, Solomon T, et al. Frequency of neurologic manifestations in COVID-19: a systematic review and meta-analysis. Neurology 2021;97:e2269–81. https://doi.org/10.1212/WNL.0000000000012930. 155. Taquet M, Geddes JR, Husain M, Luciano S, Harrison PJ. 6-month neurological and psychiatric outcomes in 236 379 survivors of COVID-19: a retrospective cohort study using electronic health records. Lancet Psychiatry 2021;8:416–27. https://doi.org/10.1016/S2215-0366(21)00084-5. 156. Cho SM, White N, Premraj L, Battaglini D, Fanning J, Suen J, et al. Neurological manifestations of COVID-19 in adults and children. Brain 2023;146:1648–61. https://doi.org/10.1093/brain/awac332. 157. Shahjouei S, Tsivgoulis G, Farahmand G, Koza E, Mowla A, Sadr AV, et al. SARS-CoV-2 and stroke characteristics: a report from the multinational COVID-19 stroke study group. Stroke 2021;52:e117–30. https://doi.org/10.1161/STROKEAHA.120.032927. 158. Merkler AE, Parikh NS, Mir S, Gupta A, Kamel H, Lin E, et al. Risk of ischemic stroke in patients with coronavirus disease 2019 (COVID-19) vs patients with influenza. JAMA Neurol 2020;77:1366–72. https://doi.org/10.1001/jamaneurol.2020.2730. 159. Katsoularis I, Fonseca-Rodríguez O, Farrington P, Lindmark K, Connolly AMF. Risk of acute myocardial infarction and ischaemic stroke following COVID-19 in Sweden: a self-controlled case series and matched cohort study. Lancet 2021;398:599–607. https://doi.org/10.1016/S0140-6736(21)00896-5. 160. Russell CD, Lone NI, Baillie JK. Comorbidities, multimorbidity and COVID-19. Nat Med 2023;29:334–43. https://doi.org/10.1038/s41591-022-02156-9. 161. World Health Organization. A clinical case definition of post COVID-19 condition by a Delphi consensus 2021. https://www.who.int/publications/i/item/WHO-2019-nCoV-Post_COVID-19_condition-Clinical_case_definition-2021.1. Accessed: November 26, 2023. 162. Xu E, Xie Y, Al-Aly Z. Long-term neurologic outcomes of COVID-19. Nat Med 2022;28:2406–15. https://doi.org/10.1038/s41591-022-02001-z. 163. Chang L, Ryan MC, Liang H, Zhang X, Cunningham E, Wang J, et al. Changes in brain activation patterns during working memory tasks in people with post-COVID condition and persistent neuropsychiatric symptoms. Neurology 2023;100:e2409–23. https://doi.org/10.1212/WNL.0000000000207309. 164. Mina Y, Enose-Akahata Y, Hammoud DA, Videckis AJ, Narpala SR, O’Connell SE, et al. Deep phenotyping of neurologic postacute sequelae of SARS-CoV-2 infection. Neurol Neuroimmunol Neuroinflamm 2023;10:e200097. https://doi.org/10.1212/NXI.0000000000200097. 165. Nalbandian A, Sehgal K, Gupta A, Madhavan MV, McGroder C, Stevens JS, et al. Post-acute COVID-19 syndrome. Nat Med 2021;27:601–15. https://doi.org/10.1038/s41591-021-01283-z. 166. Lam MHB, Wing YK, Yu MWM, Leung CM, Ma RCW, Kong APS, et al. Mental morbidities and chronic fatigue in severe acute respiratory syndrome survivors: long-term follow-up. Arch Intern Med 2009;169:2142–7. https://doi.org/10.1001/archinternmed.2009.384. 167. Aschman T, Mothes R, Heppner FL, Radbruch H. What SARS-CoV-2 does to our brains. Immunity 2022;55:1159–72. https://doi.org/10.1016/j.immuni.2022.06.013. 168. Annane D, Sharshar T. Cognitive decline after sepsis. Lancet Respir Med 2015;3:61–9. https://doi.org/10.1016/S2213-2600(14)70246-2. 169. Lamar CD, Hurley RA, Taber KH. Sepsis-associated encephalopathy: review of the neuropsychiatric manifestations and cognitive outcome. J Neuropsychiatry Clin Neurosci 2011;23:237–41. https://doi.org/10.1176/jnp.23.3.jnp237. 170. Al-Aly Z, Xie Y, Bowe B. High-dimensional characterization of post-acute sequelae of COVID-19. Nature 2021;594:259–64. https://doi.org/10.1038/s41586-021-03553-9. 171. Perez Giraldo GS, Ali ST, Kang AK, Patel TR, Budhiraja S, Gaelen JI, et al. Neurologic manifestations of long COVID differ based on acute COVID-19 severity. Ann Neurol 2023;94:146-59. https://doi.org/10.1002/ana.26649. 172. Al-Aly Z, Bowe B, Xie Y. Long COVID after breakthrough SARS-CoV-2 infection. Nat Med 2022;28:1461–7. https://doi.org/10.1038/s41591-022-01840-0. 173. Afsahi AM, Norbash AM, Syed SF, Sedaghat M, Afsahi G, Shahidi R, et al. Brain MRI findings in neurologically symptomatic COVID-19 patients: a systematic review and meta-analysis. J Neurol 2023;270:5131–54. https://doi.org/10.1007/s00415-023-11914-9. 174. Chakravarty N, Senthilnathan T, Paiola S, Gyani P, Castillo Cario S, Urena E, et al. Neurological pathophysiology of SARS-CoV-2 and pandemic potential RNA viruses: a comparative analysis. FEBS Lett 2021;595:2854–71. https://doi.org/10.1002/1873-3468.14227. 175. Hosp JA, Dressing A, Blazhenets G, Bormann T, Rau A, Schwabenland M, et al. Cognitive impairment and altered cerebral glucose metabolism in the subacute stage of COVID-19. Brain 2021;144:1263–76. https://doi.org/10.1093/brain/awab009. 176. Blazhenets G, Schroeter N, Bormann T, Thurow J, Wagner D, Frings L, et al. Slow but evident recovery from neocortical dysfunction and cognitive impairment in a series of chronic COVID-19 patients. J Nucl Med 2021;62:910–5. https://doi.org/10.2967/jnumed.121.262128. 177. Douaud G, Lee S, Alfaro-Almagro F, Arthofer C, Wang C, McCarthy P, et al. SARS-CoV-2 is associated with changes in brain structure in UK Biobank. Nature 2022;604:697–707. https://doi.org/10.1038/s41586-022-04569-5. 178. Du Y, Zhao W, Huang S, Huang C, Li C, Chen Y, et al. Gray matter thickness and subcortical nuclear volume in men after SARS-CoV-2 Omicron infection. JAMA Netw Open 2023;6:e2345626. https://doi.org/10.1001/jamanetworkopen.2023.45626. 179. Díez-Cirarda M, Yus M, Gómez-Ruiz N, Polidura C, Gil-Martínez L, Delgado-Alonso C, et al. Multimodal neuroimaging in post-COVID syndrome and correlation with cognition. Brain 2023;146:2142–52. https://doi.org/10.1093/brain/awac384. 180. Solomon IH, Singh A, Folkerth RD, Mukerji SS. What can we still learn from brain autopsies in COVID-19? Semin Neurol 2023;43:195-204. https://doi.org/10.1055/s-0043-1767716. 181. Al-Sarraj S, Troakes C, Hanley B, Osborn M, Richardson MP, Hotopf M, et al. Invited review: the spectrum of neuropathology in COVID-19. Neuropathol Appl Neurobiol 2021;47:3–16. https://doi.org/10.1111/nan.12667. 182. Lebrun L, Absil L, Remmelink M, De Mendonça R, D’Haene N, Gaspard N, et al. SARS-Cov-2 infection and neuropathological findings: a report of 18 cases and review of the literature. Acta Neuropathol Commun 2023;11:78. https://doi.org/10.1186/s40478-023-01566-1. 183. Maiese A, Manetti AC, Bosetti C, Del Duca F, La Russa R, Frati P, et al. SARS-CoV-2 and the brain: a review of the current knowledge on neuropathology in COVID-19. Brain Pathol 2021;31:e13013. https://doi.org/10.1111/bpa.13013. 184. Cosentino G, Todisco M, Hota N, Della Porta G, Morbini P, Tassorelli C, et al. Neuropathological findings from COVID-19 patients with neurological symptoms argue against a direct brain invasion of SARS-CoV-2: a critical systematic review. Eur J Neurol 2021;28:3856–65. https://doi.org/10.1111/ene.15045. 185. Thakur KT, Miller EH, Glendinning MD, Al-Dalahmah O, Banu MA, Boehme AK, et al. COVID-19 neuropathology at Columbia University Irving Medical Center/New York Presbyterian Hospital. Brain 2021;144:2696-708. https://doi.org/10.1093/brain/awab148. 186. Solomon IH, Normandin E, Bhattacharyya S, Mukerji SS, Keller K, Ali AS, et al. Neuropathological features of Covid-19. N Engl J Med 2020;383:989-92. https://doi.org/10.1056/NEJMc2019373. 187. Lou JJ, Movassaghi M, Gordy D, Olson MG, Zhang T, Khurana MS, et al. Neuropathology of COVID-19 (neuro-COVID): clinicopathological update. Free Neuropathol 2021;2:2. https://doi.org/10.17879/freeneuropathology-2021-2993. 188. Santana MF, Frank CHM, Almeida TVR, Jeronimo CMP, de Araújo Pinto RA, Martins YF, et al. Hemorrhagic and thrombotic manifestations in the central nervous system in COVID-19: a large observational study in the Brazilian Amazon with a complete autopsy series. PLoS One 2021;16:e0255950. https://doi.org/10.1371/journal.pone.0255950. 189. Hanley B, Naresh KN, Roufosse C, Nicholson AG, Weir J, Cooke GS, et al. Histopathological findings and viral tropism in UK patients with severe fatal COVID-19: a post-mortem study. Lancet Microbe 2020;1:e245–53. https://doi.org/10.1016/S2666-5247(20)30115-4. 190. Jaunmuktane Z, Mahadeva U, Green A, Sekhawat V, Barrett NA, Childs L, et al. Microvascular injury and hypoxic damage: emerging neuropathological signatures in COVID-19. Acta Neuropathol 2020;140:397–400. https://doi.org/10.1007/s00401-020-02190-2. 191. Levine A, Lee C, Fava C, Tsang F, Macneil K, Yip ST, et al. Detailed neuropathologic report of COVID-19 complicated by large intracerebral hemorrhage and periventricular lesions with macrophagic infiltrates. Free Neuropathol 2021;2:7. https://doi.org/10.17879/freeneuropathology-2021-3266. 192. El Jamal SM, Pujadas E, Ramos I, Bryce C, Grimes ZM, Amanat F, et al. Tissue-based SARS-CoV-2 detection in fatal COVID-19 infections: sustained direct viral-induced damage is not necessary to drive disease progression. Hum Pathol 2021;114:110–9. https://doi.org/10.1016/j.humpath.2021.04.012. 193. Fabbri VP, Foschini MP, Lazzarotto T, Gabrielli L, Cenacchi G, Gallo C, et al. Brain ischemic injury in COVID-19-infected patients: a series of 10 post-mortem cases. Brain Pathol 2021;31:205–10. https://doi.org/10.1111/bpa.12901. 194. Kirschenbaum D, Imbach LL, Ulrich S, Rushing EJ, Keller E, Reimann RR, et al. Inflammatory olfactory neuropathy in two patients with COVID-19. Lancet 2020;396:166. https://doi.org/10.1016/S0140-6736(20)31525-7. 195. Eschbacher KL, Larsen RA, Moyer AM, Majumdar R, Reichard RR. Neuropathological findings in COVID-19: an autopsy cohort. J Neuropathol Exp Neurol 2022;82:21–8. https://doi.org/10.1093/jnen/nlac101. 196. Al-Dalahmah O, Thakur KT, Nordvig AS, Prust ML, Roth W, Lignelli A, et al. Neuronophagia and microglial nodules in a SARS-CoV-2 patient with cerebellar hemorrhage. Acta Neuropathol Commun 2020;8:147. https://doi.org/10.1186/s40478-020-01024-2. 197. Stein JA, Kaes M, Smola S, Schulz-Schaeffer WJ. Neuropathology in COVID-19 autopsies is defined by microglial activation and lesions of the white matter with emphasis in cerebellar and brain stem areas. Front Neurol 2023;14:1229641. https://doi.org/10.3389/fneur.2023.1229641. 198. Lee MH, Perl DP, Steiner J, Pasternack N, Li W, Maric D, et al. Neurovascular injury with complement activation and inflammation in COVID-19. Brain 2022;145:2555–68. https://doi.org/10.1093/brain/awac151. 199. Stram MN, Seifert AC, Cortes E, Akyatan A, Woodoff-Leith E, Borukhov V, et al. Neuropathology of pediatric SARS-CoV-2 infection in the forensic setting: novel application of ex vivo imaging in analysis of brain microvasculature. Front Neurol 2022;13:894565. https://doi.org/10.3389/fneur.2022.894565. 200. Serrano GE, Walker JE, Tremblay C, Piras IS, Huentelman MJ, Belden CM, et al. SARS-CoV-2 brain regional detection, histopathology, gene expression, and immunomodulatory changes in decedents with COVID-19. J Neuropathol Exp Neurol 2022;81:666–95. https://doi.org/10.1093/jnen/nlac056. 201. Schurink B, Roos E, Radonic T, Barbe E, Bouman CSC, de Boer HH, et al. Viral presence and immunopathology in patients with lethal COVID-19: a prospective autopsy cohort study. Lancet Microbe 2020;1:e290–9. https://doi.org/10.1016/S2666-5247(20)30144-0. 202. Normandin E, Valizadeh N, Rudmann EA, Uddin R, Dobbins ST, MacInnis BL, et al. Neuropathological features of SARS-CoV-2 delta and Omicron variants. J Neuropathol Exp Neurol 2023;82:283–95. https://doi.org/10.1093/jnen/nlad015. 203. Wierzba-Bobrowicz T, Krajewski P, Tarka S, Acewicz A, Felczak P, Stepien T, et al. Neuropathological analysis of the brains of fifty-two patients with COVID-19. Folia Neuropathol 2021;59:219–31. https://doi.org/10.5114/fn.2021.108829. 204. Magaki S, Chen Z, Severance A, Williams CK, Diaz R, Fang C, et al. Neuropathology of microbleeds in cerebral autosomal dominant arteriopathy with subcortical infarcts and leukoencephalopathy (CADASIL). J Neuropathol Exp Neurol 2023;82:333-44. https://doi.org/10.1093/jnen/nlad004. 205. Janaway BM, Simpson JE, Hoggard N, Highley JR, Forster G, Drew D, et al. Brain haemosiderin in older people: pathological evidence for an ischaemic origin of magnetic resonance imaging (MRI) microbleeds. Neuropathol Appl Neurobiol 2014;40:258–69. https://doi.org/10.1111/nan.12062. 206. Dallaire-Théroux C, Saikali S, Richer M, Potvin O, Duchesne S. Histopathological analysis of cerebrovascular lesions associated with aging. J Neuropathol Exp Neurol 2022;81:97–105. https://doi.org/10.1093/jnen/nlab125. 207. Van Veluw SJ, Biessels GJ, Klijn CJM, Rozemuller AJM. Heterogeneous histopathology of cortical microbleeds in cerebral amyloid angiopathy. Neurology 2016;86:867–71. https://doi.org/10.1212/WNL.0000000000002419. 208. McMullen P, Smith H, Pytel P. Entrapped megakaryocytes in the microvasculature of brain tissues are not specific to COVID-19 but can be seen scross a spectrum of acute lung injuries. J Neuropathol Exp Neurol 2021;80:1078–80. https://doi.org/10.1093/jnen/nlab048. 209. Nauen DW, Hooper JE, Stewart CM, Solomon IH. Assessing brain capillaries in coronavirus disease 2019. JAMA Neurol 2021;78:760-62. https://doi.org/10.1001/jamaneurol.2021.0225. 210. Jensen MP, Le Quesne J, Officer-Jones L, Teodòsio A, Thaventhiran J, Ficken C, et al. Neuropathological findings in two patients with fatal COVID-19. Neuropathol Appl Neurobiol 2021;47:17-25. https://doi.org/10.1111/nan.12662. 211. Reichard RR, Kashani KB, Boire NA, Constantopoulos E, Guo Y, Lucchinetti CF. Neuropathology of COVID-19: a spectrum of vascular and acute disseminated encephalomyelitis (ADEM)-like pathology. Acta Neuropathol 2020;140:1–6. https://doi.org/10.1007/s00401-020-02166-2. 212. Walker JM, Gilbert AR, Bieniek KF, Richardson TE. COVID-19 patients with CNS complications and neuropathologic features of acute disseminated encephalomyelitis and acute hemorrhagic leukoencephalopathy. J Neuropathol Exp Neurol 2021;80:628–31. https://doi.org/10.1093/jnen/nlab036. 213. Manzano GS, McEntire CRS, Martinez-Lage M, Mateen FJ, Hutto SK. Acute disseminated encephalomyelitis and acute hemorrhagic leukoencephalitis following COVID-19: systematic review and meta-synthesis. Neurol Neuroimmunol Neuroinflammation 2021;8:e1080. https://doi.org/10.1212/NXI.0000000000001080. 214. Kantonen J, Mahzabin S, Mäyränpää MI, Tynninen O, Paetau A, Andersson N, et al. Neuropathologic features of four autopsied COVID-19 patients. Brain Pathol 2020;30:1012–6. https://doi.org/10.1111/bpa.12889. 215. Lee MH, Perl DP, Nair G, Li W, Maric D, Murray H, et al. Microvascular injury in the brains of patients with Covid-19. N Engl J Med 2021;384:481–3. https://doi.org/10.1056/NEJMc2033369. 216. Mavrikaki M, Lee JD, Solomon IH, Slack FJ. Severe COVID-19 is associated with molecular signatures of aging in the human brain. Nat Aging 2022;2:1130–7. https://doi.org/10.1038/s43587-022-00321-w. 217. Fullard JF, Lee HC, Voloudakis G, Suo S, Javidfar B, Shao Z, et al. Single-nucleus transcriptome analysis of human brain immune response in patients with severe COVID-19. Genome Med 2021;13:118. https://doi.org/10.1186/s13073-021-00933-8. 218. Mukherjee A, Peterson JEG, Sandberg G, Takei H, Adesina A, Goodman JC, et al. Central nervous system pathology in fatal swine-origin influenza A H1N1 virus infection in patients with and without neurological symptoms: an autopsy study of 15 cases. Acta Neuropathol 2011;122:371–3. https://doi.org/10.1007/s00401-011-0854-7. 219. Gu J, Korteweg C. Pathology and pathogenesis of severe acute respiratory syndrome. Am J Pathol 2007;170:1136–47. https://doi.org/10.2353/ajpath.2007.061088. 220. Gu J, Gong E, Zhang B, Zheng J, Gao Z, Zhong Y, et al. Multiple organ infection and the pathogenesis of SARS. J Exp Med 2005;202:415–24. https://doi.org/10.1084/jem.20050828. 221. Xu J, Zhong S, Liu J, Li L, Li Y, Wu X, et al. Detection of severe acute respiratory syndrome coronavirus in the brain: potential role of the chemokine mig in pathogenesis. Clin Infect Dis 2005;41:1089–96. https://doi.org/10.1086/444461. 222. Lau KK, Yu WC, Chu CM, Lau ST, Sheng B, Yuen KY. Possible central nervous system infection by SARS coronavirus. Emerg Infect Dis 2004;10:342–4. https://doi.org/10.3201/eid1002.030638. 223. Hung EC, Chim SS, Chan PK, Tong YK, Ng EK, Chiu RW, et al. Detection of SARS coronavirus RNA in the cerebrospinal fluid of a patient with severe acute respiratory syndrome. Clin Chem 2003;49:2108–9. https://doi.org/10.1373/clinchem.2003.025437. 224. Ding Y, He L, Zhang Q, Huang Z, Che X, Hou J, et al. Organ distribution of severe acute respiratory syndrome (SARS) associated coronavirus (SARS-CoV) in SARS patients: implications for pathogenesis virus transmission pathways. J Pathol 2004;203:622–30. https://doi.org/10.1002/path.1560. 225. Bhatnagar J, Gary J, Reagan-Steiner S, Estetter LB, Tong S, Tao Y, et al. Evidence of severe acute respiratory syndrome coronavirus 2 replication and tropism in the lungs, airways, and vascular endothelium of patients with fatal coronavirus disease 2019: an autopsy case series. J Infect Dis 2021;223:752–64. https://doi.org/10.1093/infdis/jiab039. 226. Remmelink M, De Mendonça R, D’Haene N, De Clercq S, Verocq C, Lebrun L, et al. Unspecific post-mortem findings despite multiorgan viral spread in COVID-19 patients. Crit Care 2020;24:495. https://doi.org/10.1186/s13054-020-03218-5. 227. Fabbri VP, Riefolo M, Lazzarotto T, Gabrielli L, Cenacchi G, Gallo C, et al. COVID-19 and the brain: the neuropathological Italian experience on 33 adult autopsies. Biomolecules 2022;12:629. https://doi.org/10.3390/biom12050629. 228. Boluda S, Mokhtari K, Mégarbane B, Annane D, Mathon B, Cao A, et al. Golgi localization of SARS-CoV-2 spike protein and interaction with furin in cerebral COVID-19 microangiopathy: a clue to the central nervous system involvement? Free Neuropathol 2023;4:4-1. https://doi.org/10.17879/freeneuropathology-2023-4584. 229. Song E, Zhang C, Israelow B, Lu-Culligan A, Prado AV, Skriabine S, et al. Neuroinvasion of SARS-CoV-2 in human and mouse brain. J Exp Med 2021;218:e20202135. https://doi.org/10.1084/JEM.20202135. 230. Matschke J, Lütgehetmann M, Hagel C, Sperhake JP, Schröder AS, Edler C, et al. Neuropathology of patients with COVID-19 in Germany: a post-mortem case series. Lancet Neurol 2020;19:919–29. https://doi.org/10.1016/S1474-4422(20)30308-2. 231. Meinhardt J, Radke J, Dittmayer C, Franz J, Thomas C, Mothes R, et al. Olfactory transmucosal SARS-CoV-2 invasion as a port of central nervous system entry in individuals with COVID-19. Nat Neurosci 2021;24:168–75. https://doi.org/10.1038/s41593-020-00758-5. 232. Bauer L, Laksono BM, de Vrij FMS, Kushner SA, Harschnitz O, van Riel D. The neuroinvasiveness, neurotropism, and neurovirulence of SARS-CoV-2. Trends Neurosci 2022;45:358–68. https://doi.org/10.1016/j.tins.2022.02.006. 233. de Melo GD, Lazarini F, Levallois S, Hautefort C, Michel V, Larrous F, et al. COVID-19-related anosmia is associated with viral persistence and inflammation in human olfactory epithelium and brain infection in hamsters. Sci Transl Med 2021;13:eabf8396. https://doi.org/10.1126/scitranslmed.abf8396. 234. Durrant DM, Ghosh S, Klein RS. The olfactory bulb: an immunosensory effector organ during neurotropic viral infections. ACS Chem Neurosci 2016;7:464–9. https://doi.org/10.1021/acschemneuro.6b00043. 235. Lopez G, Tonello C, Osipova G, Carsana L, Biasin M, Cappelletti G, et al. Olfactory bulb SARS-CoV-2 infection is not paralleled by the presence of virus in other central nervous system areas. Neuropathol Appl Neurobiol 2022;48:e12752. https://doi.org/10.1111/nan.12752. 236. Krasemann S, Dittmayer C, von Stillfried S, Meinhardt J, Heinrich F, Hartmann K, et al. Assessing and improving the validity of COVID-19 autopsy studies - a multicentre approach to establish essential standards for immunohistochemical and ultrastructural analyses. EBioMedicine 2022;83:104193. https://doi.org/10.1016/j.ebiom.2022.104193. 237. Jarius S, Pache F, Körtvelyessy P, Jelčić I, Stettner M, Franciotta D, et al. Cerebrospinal fluid findings in COVID-19: a multicenter study of 150 lumbar punctures in 127 patients. J Neuroinflammation 2022;19:19. https://doi.org/10.1186/s12974-021-02339-0. 238. Lersy F, Benotmane I, Helms J, Collange O, Schenck M, Brisset JC, et al. Cerebrospinal fluid features in patients with coronavirus disease 2019 and neurological manifestations: correlation with brain magnetic resonance imaging findings in 58 patients. J Infect Dis 2021;223:600–9. https://doi.org/10.1093/infdis/jiaa745. 239. Destras G, Bal A, Escuret V, Morfin F, Lina B, Josset L. Systematic SARS-CoV-2 screening in cerebrospinal fluid during the COVID-19 pandemic. Lancet Microbe 2020;1:e149. https://doi.org/10.1016/S2666-5247(20)30066-5. 240. Edén A, Grahn A, Bremell D, Aghvanyan A, Bathala P, Fuchs D, et al. Viral antigen and inflammatory biomarkers in cerebrospinal fluid in patients with COVID-19 infection and neurologic symptoms compared with control participants without infection or neurologic symptoms. JAMA Netw Open 2022;5:e2213253. https://doi.org/10.1001/jamanetworkopen.2022.13253. 241. Hopfer H, Herzig MC, Gosert R, Menter T, Hench J, Tzankov A, et al. Hunting coronavirus by transmission electron microscopy – a guide to SARS-CoV-2-associated ultrastructural pathology in COVID-19 tissues. Histopathology 2021;78:358–70. https://doi.org/10.1111/his.14264. 242. Goldsmith CS, Miller SE, Martines RB, Bullock HA, Zaki SR. Electron microscopy of SARS-CoV-2: a challenging task. Lancet 2020;395:e99. https://doi.org/10.1016/S0140-6736(20)31188-0 243. Bullock HA, Goldsmith CS, Zaki SR, Martines RB, Miller SE. Difficulties in differentiating coronaviruses from subcellular structures in human tissues by electron microscopy. Emerg Infect Dis 2021;27:1023–31. https://doi.org/10.3201/eid2704.204337. 244. Xekardaki A, Santos M, Hof P, Kövari E, Bouras C, Giannakopoulos P. Neuropathological substrates and structural changes in late-life depression: the impact of vascular burden. Acta Neuropathol 2012;124:453–64. https://doi.org/10.1007/s00401-012-1021-5. 245. Jellinger KA. The heterogeneity of late-life depression and its pathobiology: a brain network dysfunction disorder. J Neural Transm 2023;130:1057–76. https://doi.org/10.1007/s00702-023-02648-z. 246. Reiken S, Sittenfeld L, Dridi H, Liu Y, Liu X, Marks AR. Alzheimer’s-like signaling in brains of COVID-19 patients. Alzheimer’s Dement 2022;18:955–65. https://doi.org/10.1002/alz.12558. 247. Nelson PT, Head E, Schmitt FA, Davis PR, Neltner JH, Jicha GA, et al. Alzheimer’s disease is not "brain aging": Neuropathological, genetic, and epidemiological human studies. Acta Neuropathol 2011;121:571–87. https://doi.org/10.1007/s00401-011-0826-y. 248. Horvath S, Levine AJ. HIV-1 infection accelerates age according to the epigenetic clock. J Infect Dis 2015;212:1563–73. https://doi.org/10.1093/infdis/jiv277. 249. Khanlou N, Moore DJ, Chana G, Cherner M, Lazzaretto D, Dawes S, et al. Increased frequency of alpha-synuclein in the substantia nigra in human immunodeficiency virus infection. J Neurovirol 2009;15:131–8. https://doi.org/10.1080/13550280802578075. 250. Levine A, Sacktor N, Becker JT. Studying the neuropsychological sequelae of SARS-CoV-2: lessons learned from 35 years of neuroHIV research. J Neurovirol 2020;26:809–23. https://doi.org/10.1007/s13365-020-00897-2. 251. Voruz P, Jacot De Alcântara I, Nuber-Champier A, Cionca A, Allali G, Benzakour L, et al. Frequency of abnormally low neuropsychological scores in post-COVID-19 syndrome: the Geneva COVID-COG Cohort. Arch Clin Neuropsychol 2023;38:1–11. https://doi.org/10.1093/arclin/acac068. 252. Lynch S, Ferrando SJ, Dornbush R, Shahar S, Smiley A, Klepacz L. Screening for brain fog: Is the montreal cognitive assessment an effective screening tool for neurocognitive complaints post-COVID-19? Gen Hosp Psychiatry 2022;78:80–6. https://doi.org/10.1016/j.genhosppsych.2022.07.013. 253. Lou J, Movassaghi M, Gordy D, Olson M, Singer E, Magaki S, et al. Data elements and tissue procurement for COVID-19 brain banking. J Neuropathol Exp Neurol 2021;80:579–80. 254. Morgello S, Gelman BB, Kozlowski PB, Vinters HV, Masliah E, Cornford M, et al. The National NeuroAIDS Tissue Consortium: a new paradigm in brain banking with an emphasis on infectious disease. 2001;27:326-35. https://doi.org/10.1046/j.0305-1846.2001.00334.x. 255. Hooper JE. Rapid autopsy programs and research support: the pre- and post-COVID-19 environments. AJSP Rev Rep 2021;26:100–7. PMC7954201. 256. Zhu Y, Wang L, Yin Y, Yang E. Systematic analysis of gene expression patterns associated with postmortem interval in human tissues. Sci Rep 2017;7:5435. https://doi.org/10.1038/s41598-017-05882-0. 257. Skrobot OA, Attems J, Esiri M, Hortobágyi T, Ironside JW, Kalaria RN, et al. Vascular cognitive impairment neuropathology guidelines (VCING): the contribution of cerebrovascular pathology to cognitive impairment. Brain 2016;139:2957-69. https://doi.org/10.1093/brain/aww214.
Copyright: © 2024 The author(s). This is an open access article distributed under the terms of the Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited, a link to the Creative Commons license is provided, and any changes are indicated. The Creative Commons Public Domain Dedication waiver (https://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated. |