|
|
|
Free Neuropathology 4:3 (2023) |
|
Review |
|
Multiple sclerosis: 2023 update |
|
Tanja Kuhlmann1,2, Jack Antel2 |
|
1 Institute of Neuropathology, University Hospital Münster, 48149 Münster, Germany |
|
Corresponding author: |
|
Submitted: 02 February 2023 |
|
Keywords: Multiple sclerosis, EBV, Remyelination, Slowly expanding lesions, Spatial transcriptomics |
|
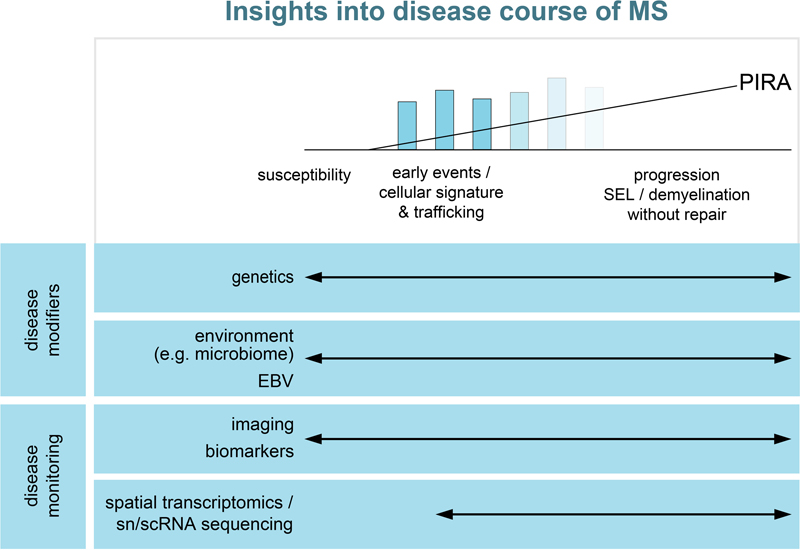
Abstract Multiple sclerosis (MS) is the most frequent inflammatory and demyelinating disease of the Central Nervous System (CNS). Significant progress has been made during recent years in preventing relapses by using systemic immunomodulatory or immunosuppressive therapies. However, the limited effectiveness of such therapies for controlling the progressive disease course indicates there is a continuous disease progression independent of relapse activity which may start very early during the disease course. Dissecting the underlying mechanisms and developing therapies for preventing or stopping this disease progression represent, currently, the biggest challenges in the field of MS. Here, we summarize publications of 2022 which provide insight into susceptibility to MS, the basis of disease progression and features of relatively recently recognized distinct forms of inflammatory/demyelinating disorders of the CNS, such as myelin oligodendrocyte glycoprotein antibody-associated disease (MOGAD). Introduction Multiple sclerosis (MS) is the most frequent inflammatory demyelinating disease of the Central Nervous System (CNS) and a leading cause for permanent neurological disability in young adults. The variability in disease course amongst cases is well known. Significant progress has been made during recent years in using systemic immune-system directed therapies to prevent relapses that characterize the initial disease course. However, the limited effectiveness of such therapies for the later-recognized progressive disease course indicates there is a continuous disease progression independent of relapse activity which may start very early during this condition 1,2. We select articles published in 2022 that provide insight into susceptibility to MS, the basis of disease progression, and features of relatively recently recognized distinct forms of inflammatory/demyelinating disorders of the CNS (Figure 1). MS has a strong hereditary component, but also environmental factors play an important role, as shown in studies of monozygotic twins discordant for the disease. Several articles published in 2022 support that Epstein-Barr virus (EBV), which has long been implicated in MS, plays an important role in the development of MS. This raises the tantalizing possibility that vaccination against EBV may eliminate MS. Another topic, which continues to be heavily discussed in the MS field, is the impact of the microbiome, not only in the gut but also in the lung, on MS disease course. Potential mechanisms driving disease progression include persistent focal inflammation resulting in slowly expanding lesions, meningeal inflammation causing cortical demyelination and neuronal injury, and inflammation induced diffuse changes in white matter (WM) as well as loss of compensatory mechanism, such as remyelination and brain plasticity 3. We summarize several pathological and imaging studies, which aim to dissect the underlying mechanisms driving disease progression. However, these mechanisms most likely vary over time, adding another layer of complexity for the successful development of new treatment strategies to prevent or stop disease progression. Furthermore, individual patient specific factors may modulate MS pathology significantly. This is supported by results from a single nuclei RNA sequencing (snRNAseq) study of human MS tissue samples which reveals that the variability in gene expression between patients is higher than between lesions 4. Based on findings from animal studies, it is assumed that promotion of remyelination can prevent disease progression. However, the evidence that remyelination in MS is neuroprotective is relatively sparse. In 2022, an imaging study provides first evidence that lack of remyelination in MS is associated with increased brain atrophy, a maker of neurodegeneration 5.
Figure 1: Insights into disease course of MS. We selected articles published in 2022 providing insights into the susceptibility to MS, potential modulators of the disease course and new technologies to monitor the disease or to dissect underlying pathways. Finally, we discuss the most recent findings in myelin oligodendrocyte glycoprotein antibody-associated disease (MOGAD), a relatively recently newly defined inflammatory and demyelinating disease recognized by the presence of anti-myelin oligodendrocyte glycoprotein (MOG) antibodies in the serum. (I) MS susceptibility and disease course – population based studies Role of EBV in susceptibility to MS Epidemiologic based studies in 2022 bolstered the evidence for EBV infection being a necessary albeit, not sufficient trigger, for development of MS. Bjornevik et al. examined the records of a cohort of more than 10 million active US military personnel in which 801 of 955 cases that developed MS could be evaluated with regard to EBV serology 6. All but one were serologically positive at the time of diagnosis. Although 35 had initially been serologically negative, 34 converted prior to diagnosis. Seroconversion rate was 57% in the non-MS cohort. No enhanced conversion rate was found for other viruses; cytomegalovirus (CMV) conversion may have been protective. In a retrospective cohort study of 2,576,011 individuals born in Denmark between 1971 and 2018, Rostgaard et al. found that having a sibling 0–2 years younger reduces the probability of MS by 25% compared to having no siblings. Furthermore, having a second sibling 6–8 years younger reduces this probability by 30% 7. They concluded that having younger siblings reduces the risk of infectious mononucleosis and the risk of MS in a mirror-like manner, suggesting that the reduction in MS risk results from earlier primary EBV infection. The basis whereby EBV infection links to disease susceptibility and/or disease course remains under investigation. EBV is known to infect B cells and these cells have been detected in MS lesions in some studies, but still require unequivocal confirmation. B cells are implicated in producing factors that contribute to underlying tissue injury. Support for the postulate that EBV specific humoral and/or cellular immune mediators contribute to the disease pathology is supported by the findings by Lanz et al., regarding cross-reactivity between EBV and neural tissue components 8. They demonstrated high affinity molecular mimicry between the EBV transcription factor Epstein–Barr nuclear antigen 1 (EBNA- 1) and the CNS protein GlialCAM, as well as presence of cross reactive antibodies in the cerebrospinal fluid (CSF) of MS patients. Interestingly, these cross-reactive antibodies bound to rodent oligodendrocytes and astrocytes, glia limitans and, likely, perivascular glial cells in the hippocampus and brainstem, a pattern reminiscent of the expression pattern of aquaporin-4 (AQP4). Anti-AQP4 antibodies are the hallmark of neuromyelitis optica (NMO), another inflammatory demyelinating disease with pathological features distinct from MS 9,10. Further support for the potential relevance of EBV specific humoral and/or cellular immune mediators in MS pathogenesis is the study by Schneider-Hohendorf et al. conducted on MS disease discordant monozygotic twins, showing a broader EBV-specific T cell receptor (TCR) repertoire in the affected one of the pair 11. These combined population and laboratory based studies raise the issue of potential benefit of EBV directed vaccines in the very young population. However, one need consider the potential risk of such vaccination, including inducing a neural-target directed cross reactive immune response and persistent modulation of overall immune-regulatory mechanisms which may have protective purposes. Microbiome impacts on disease susceptibility and course in MS Having identified that an array of environmental factors contribute to the susceptibility and course of MS, the challenge remains to understand the underlying mechanisms for such effects. Further research is needed to find out how these factors can act via modulating the systemic immune system to acquire pro-inflammatory properties. To dissect the influence of genetic predisposition versus environmental factors on shaping the immune system, Ingelfinger et al. applied a combination of antibody-based and single-cell technologies to define the peripheral immune signatures of 61 monozygotic (MZ) twin pairs discordant for MS (+ healthy MZ twin and dizygotic twin pairs) 12. They could discern both genetically (CD25 expression by naïve helper T cells) and environmentally (helper T cells with a dysregulated CD25-IL-2 axis) determined features of an MS-associated immune signature. The microbiome serves as a major interaction of a host with the environment. The gut is long recognized as a major resident niche for the immune system, being a major site of host immune interaction with the enormous population of resident microbes. The results of the International Multiple Sclerosis Microbiome Study (iMSMS), derived from an analysis of the gut microbiome of 576 MS patients (36% untreated) and genetically unrelated household healthy controls (1,152 total subjects), indicated that the phytate degradation pathway was over-represented in untreated MS, while pyruvate-producing carbohydrate metabolism pathways were significantly reduced 13. Microbiome composition, function, and derived metabolites were all modified in response to disease-modifying treatments. As pointed out by Hosang et al., the lung (airway) also serves as a niche for immune cells supporting their long-term survival and maturation into migration-competent effector T cells 14. Smoking is a recognized risk factor for the development of MS. Hosang et al. demonstrated the potential for the interaction of this niche with the environment to influence the susceptibility to autoimmune neurologic disease, using the experimental autoimmune encephalomyelitis (EAE) model. Treating animals with antibiotics (neomycin) shifted the microbiota towards lipopolysaccharide (LPS)-enriched phyla without an effect on the gut microbiome. The underlying mechanism of effect implicated was that LPS would usually induce a type 1 interferon response in microglia and reduce immune activation within the CNS. However, LPS is also used to induce a pro-inflammatory response in human microglia 15. The above studies indicate the potential for the microbiome in multiple niches to impact on MS throughout its disease course. Microbiome influences on adaptive immune cells would impact on their initial activation and migration into the CNS as part of new lesion formation. Modulation of the endogenous glia cells populations (astrocytes, microglia) by products derived from the microbiome would impact on the extent to which these glial cells participate in tissue injury and repair processes throughout the MS disease course. Progression independent of relapse activity The natural history of multiple sclerosis has been considered in terms of relapsing and progressive categories; the latter could occur in those with previous clinical relapses (secondary progressive (SP)) or not (primary progressive (PP)). Specific criteria have been defined for diagnosis of either relapsing or PP forms of the disease that now include paraclinical measures (CSF, imaging) as well as clinical features 16. Progression needs not be sustained in either SP or PP forms resulting in use of the term “confirmed progression” (over 3-6 months) in clinical trials. More recent focus is whether progressive forms are associated with ongoing disease activity, as defined by imaging or clinical activity, introducing the term progression independent of relapse activity (PIRA) 1,2,17. This designation has acquired increased significance in that regulatory approval for agents in SP MS has been denied for cases lacking of documented activity. Lublin et al. used the Novartis-Oxford multiple sclerosis (NO.MS) data pool, spanning all multiple sclerosis phenotypes and paediatric multiple sclerosis, to evaluate ∼200,000 Expanded Disability Status Scale (EDSS) transitions from >27,000 patients with ≤15 years follow-up 18. Their conclusions were that although relapses contribute to the accumulation of disability, primarily early in multiple sclerosis, PIRA started early in the disease process, occurred in all phenotypes and became the principal driver of disability accumulation in the progressive phase of the disease. Tur et al. could show that PIRA, after a first demyelinating event, was not uncommon and suggested an unfavorable long-term prognosis, especially if it occurs early in the disease course 19. Focal inflammation as driver of disease progression The observation that disease progression is driven by mechanisms other than peripherally-driven acute focal inflammatory lesions, raise the issue of the pathogenic mechanisms responsible for progression. This has become a hot topic for both histopathologic and serial neuroimaging studies, with a challenge of how to reconcile these two approaches. Both have implicated pathogenic processes in the white matter, cortical grey, periventricular, and spinal cord regions of the CNS. As regards focal white matter lesions, Beynon et al. examined chronic lesion activity (CLA) ( assessed by T1-hypointense lesion volume increase within baseline T2-non-enhancing lesions identified as either slowly expanding (SELs) or not slowly expanding (non-SELs)) in a 108 week phase 3 clinical trial of natalizumab (ASCEND) in SP MS patients 20. They found that CLA in both SELs and non-SELs was greater in patients with SPMS and confirmed disability progression than in those with no progression. SEL prevalence was lower in the absence of acute lesion activity (ALA) (5% vs. 19% in subjects with ALA), while CLA remained associated with disability progression. In view of the association of ALA with SEL prevalence, the authors suggest that the effect of natalizumab on CLA may be related to its effect on ALA and the suppression of ALA in mixed active/inactive lesions. Further studies are required to understand whether and to which extent the established immunosuppressive and immunomodulatory MS treatments target SEL (with or without paramagnetic rims). (II) MS susceptibility and disease course – Tissue based studies Meningeal inflammation correlates with lesion activity The meninges consists of the dura mater and the leptomeninges comprising pia mater and arachnoid. During recent years, a number of papers have elucidated the relationship between leptomeningeal inflammation and subpial cortical lesions 21-23. Subpial lesions are more frequently found in close proximity to meningeal infiltrates and, subpial demyelination is associated with neuronal, oligodendroglial and astrocytic injury and loss; cortical pathology is now considered as a driver of disease progression 22. Subpial lesions are characterized by reduced extent of grey matter (GM) inflammation compared to white matter lesions; therefore, the question occurs whether additional mechanisms may contribute to the development of these lesions. An obvious possibility would be that soluble factors secreted by the inflammatory cells contribute to subpial demyelination and cellular injury. Earlier pathological studies could not identify a correlation between meningeal inflammation and number of WM lesions 23,24. Ahmed et al. addressed the question whether meningeal infiltrates correlate with WM lesion activity 25. They analyzed the density of meningeal lymphocytes and their topographical association with grey (cortical) and white matter lesions in 27 patients with progressive MS and nine controls. Numbers of T and B cells per unit length of meninges were significantly increased in MS compared to controls and number of meningeal lymphocytes correlated with extent of subpial demyelination, confirming earlier studies 22,23,24,26. Patients with high meningeal lymphocytic counts had more cortical lesions compared to patients with low lymphocytic counts, but no difference in the percentage of subpial, leukocortical or intracortical lesions. Importantly, patients with a high density of meningeal lymphocytic infiltrates had a higher proportion of active and mixed lesions and a lower percentage of inactive or remyelinated lesions than patients with a low density of meningeal infiltrates. Interestingly, no correlation between meningeal myeloid infiltrates and subpial demyelination or WM lesion activity was detected. The authors speculated that the correlation of the extent of meningeal inflammation and the presence of higher proportions of active and mixed lesions may represent two independent pathogenic mechanisms which reflect an overall higher disease activity. In line with this observation are studies which demonstrated a correlation between numbers of leptomeningeal B cells and perivascular T and B cells in WM or brain stem lesions 27,28. Cellular trafficking to and from the meninges The bi-directional communication between the systemic and CNS compartments is a central process contributing to the disease course of multiple sclerosis. To be considered are the connections whereby systemic immune constituents can access the CNS and how materials released within the CNS are cleared from this compartment and transferred back to the systemic compartment. In context of immune-brain interactions, entry via the blood-brain barrier (BBB) has been the classic pathway. Blocking immune cell passage with adhesion molecule directed inhibitors or antibodies is an effective therapy in the EAE model and translates into clinical therapy (natalizumab is an approved therapy for relapsing MS). A further pathway involves blood-CSF trafficking that underlies immune seeding within the meninges. The observed pathologic changes in MS in the subpial grey matter and periventricular regions support the impact of immune constituents from the CSF-meninges acting on the brain parenchyma. Drieu et al., taking advantage of the capacity to pharmacologically or genetically delete selective cell populations in mice, showed that populations of perivascular and leptomeningeal macrophages, referred to as parenchymal border macrophages (PBMs), regulate arterial motion that drives CSF flow 29. The CNS 'waste clearance' system is defined as the glymphatic system. Carotenuto et al. used an array of different magnetic resonance imaging (MRI) techniques to identify dysfunction of the glymphatic system function in multiple sclerosis and found an association with clinical disability, disease course, demyelination and neurodegeneration 30. Transport of neural tissue breakdown products, specifically myelin constituents, back to regional lymph nodes has previously been documented by showing their presence in cervical lymph nodes of MS cases 31.The detection of lymphatic dural vessels, which connect CSF and draining cervical lymph nodes, raises the question of the relevance of these vessels and dural inflammation for autoimmune disease such as multiple sclerosis 32. Merlini et al. have studied the functional role of dural and leptomeningeal inflammation in EAE using intravital microscopy to address this question 32. They could show that the leptomeninges were highly inflamed in acute and chronic EAE, whereas the dura was only minimal affected. T cells adhered more weakly, antigen presentation was less efficient and activation of autoreactive T cells was lower in the dura compared to the leptomeninges suggesting that the dura plays only a minor role in autoimmune inflammation. These observations were matched by findings in MS brains. Histological studies analyzing the extent of inflammation in leptomeninges compared to the dura observed significant T cell infiltrates only in the leptomeninges, but not in the dura. Diffuse white mater inflammation and it consequences for the myelin-axon unit The pathogenic mechanisms driving disease progression independent of new lesions are, so far, only incompletely understood. Potential mechanisms include the expansion of focal lesions, as described above, but also diffuse changes in normal appearing white matter in MS. In their study, van den Bosch et al. elucidate the consequences of diffuse microglia activation and low-level T cell infiltration for axon-myelin units in the normal appearing white matter (NAWM), using high-resolution immunohistochemistry (IHC) and transmission electron microscopy (TEM) 33. They compared the ultrastructural characteristics of the axon-myelin unit in non-lesional optic nerves from 8 people with MS and 8 controls. In the optic nerves of MS patients, they observed, as expected, more activated and phagocytic microglia and more T cells than in controls. They analyzed the structure of the nodes of Ranvier and found an elongation of paranodes and juxtaparanodes, as well as an increased overlap between paranodal and juxtaparanodal regions, confirming the results of earlier studies 34,35. When analyzing the structure of the myelin sheath, they found a decrease in the g-ratio compared to controls, suggesting a thicker myelin sheath in MS optic nerves. However, more detailed analyses demonstrated a loss of myelin compaction as the underlying cause for the decreased g-ratio. TEM analyses revealed a higher percentage of axons containing mitochondria in patients with MS, which was confirmed by high-resolution IHC. The overlap of paranodes and juxtaparanodes, less compacted myelin and mitochondrial frequency in axons correlated with the number of activated microglia. These findings suggest that activated microglia contribute to nodal and myelin disorganization in NAWM in MS patients, which may result in a higher axonal energy demand and increased mitochondrial numbers to compensate it. Further studies are required to determine whether the same changes occur throughout the CNS in patients with MS, when these changes start and whether these pathological changes correlate with PIRA. (III) MS susceptibility and disease course - new technologies In vivo imaging to assess consequences of demyelination and presence of re-myelination in MS Emerging concepts suggest that a combination of persisting focal and diffuse inflammation within the CNS and a gradual failure of compensatory mechanisms, including remyelination and brain plasticity, result in disease progression. In demyelinating animal models, remyelination reduces axonal as well as neuronal damage and is associated with clinical recovery 36-38. Therefore, promotion of remyelination appears to be a promising new treatment approach in MS to prevent disease progression. However, despite increasing knowledge regarding the molecular mechanisms regulating de- and remyelination in animal models, the development of new remyelination promoting drugs is still a challenge. To date, clinical trials testing experimentally identified drugs have had limited success. The failure of these clinical trials might be at least partly explained by our still limited knowledge about the natural course and clinical effects of remyelination in MS patients. Tonietto et al. combined voxel based positron emission tomography (PET) and magnetization transfer MRI (MTR) approaches to study the influence of lesion location on myelin repair and the correlation between remyelination and grey matter atrophy 5. At baseline, the probability of demyelinated voxels was highest close to the ventricle and, increasing distance from the ventricle was associated with an increased myelin content in lesions. They also reported that the probability of a demyelinated voxel to remyelinate in the follow-up period correlated with the distance from the ventricle. These findings are in line with histopathological studies, observing a lower extent of remyelination in periventricular compared to subcortical lesions 39,40. Interestingly, Tonietto et al. also observed that failure of remyelination was associated with lower thalamic volume, an imaging marker of neurodegeneration 5. Additionally, they found a weak association between periventricular remyelination failure and regional cortical atrophy depending on the number of cortex-derived tracks passing through periventricular white matter. These finding suggest that lack of periventricular remyelination contributes to cortical damage in MS. However, the small sample size and the relatively short follow-up are limitations of these findings. Further studies are required to determine the relationship between remyelination (failure) and brain atrophy. Arnold et al. reported that Siponimod therapy (EXPND trial) improved magnetic transfer ratios (MTR), a measure of myelination within new lesions and across grey and white matter 41. This was associated with reduced loss of cortical grey matter, thalamic, and total brain volume, again suggesting a beneficial effect of remyelination for brain atrophy. As mentioned, the challenge remains to distinguish therapeutic effects acting via impact on inflammation from neuroprotective and repair promoting effects. To date, no therapy has received regulatory approval based entirely on the latter. A challenge remains how to use advances in imaging (and other para-clinical measures (electrophysiology, biomarkers)) to rapidly predict meaningful efficacy of therapies aimed at the latter, as so successfully done for immune-therapies for relapsing MS. New imaging techniques may help to differentiate repair and injury mechanisms. Rahmandazeh et al. combined cross-sectional, longitudinal and post-mortem histopathology-imaging validation studies to evaluate, for the first time, the usefulness of quantitative susceptibility mapping (QSM) for the identification of different MS lesion types, including remyelinated lesions 42. QSM quantifies the distribution of magnetic susceptibility in tissues and is sensitive to myelin content and iron accumulation in the brain 43,44. Due to its ability to detect iron, it has been used in the past for the detection of paramagnetic rim lesions (PRLs), which histopathologically, at least partly, correspond to mixed active/inactive lesions (= chronic active lesions). In this cross-sectional study, the authors analyzed 1,621 lesions and classified them in the QSM maps as either isointense (29 %), hypointense (4 %), hyperintense (52 %), lesions with a hypointense rim (1 %) and PRLs (13 %). The QSM histopathological correlation study revealed that iso/hypointense lesions mostly corresponded to remyelinated lesions, hyperintense lesions to inactive lesions and PRLs mostly to mixed active/inactive lesions. However, one caveat is that hypo-intensity in QSM indicates complete remyelination in the absence of ongoing inflammation; therefore, it appears less suitable for the detection of ongoing remyelination during the first five to six months after lesion formation 45. Further studies comparing different imaging technologies with histopathological findings are required to evaluate the advantages and limitations of these approaches in the follow-up of remyelination in people with MS. Novel molecular technologies to dissect MS pathogenesis in human tissue sections A number of studies so far have analyzed human MS tissue samples by snRNAseq 46-48. However, these studies included only a relatively small number of tissue samples from few patients. Macnair et al. analyzed in their not yet peer-reviewed study the largest cohort of MS and control tissue samples by snRNAseq 4. They analysed 740,000 snRNAseq profiles from 165 samples including WM lesions (n = 62), normal appearing WM (n = 17), GM lesions (n = 39) and normal appearing GM (n = 16) from 55 MS patients, as well as, 15 WM and 16 GM tissue samples from 28 controls. They reached a median sequencing depth of 4,194 nuclei/sample, 3,154 reads/nucleus and 1,702 genes per nucleus. Unsurprisingly, the authors observed highly cell-type specific changes in gene expression in WM and GM lesions in MS. Interestingly, cell-type specific changes were largely shared across lesions, and patient identity was a much stronger driver of variability in cell type-specific expression for many genes than lesion type. Nevertheless, since the composition of MS lesions is heterogeneous and the authors did not dissect specific lesion areas, for example the rim of mixed lesions, more specific signatures may have been overlooked. The authors observed an enrichment of MS risk genes differentially expressed in immune cells, but also in neurons, astrocytes and oligodendrocytes, supporting the notion that not only immune cells but also neural cells play a role for MS disease risk and progression. They also found an enrichment of different oligodendroglial subpopulations in lesions and NAWM in MS compared to control WM; however, again donor ID was a stronger driver of variability than lesion type. They identified three different oligodendroglial patterns. The first pattern was similar to controls, the second was characterized by a high proportion of an oligodendroglial subtype upregulating cellular stress genes and, the third pattern showed high proportions of early oligodendrocytes and reduced levels of more mature oligodendrocytes, suggestive of an arrest of oligodendroglial maturation or differentiation. These data suggest that patients may respond differently to potential remyelination promoting drugs. The challenge for the future will be to develop measure to identify patients who will especially benefit from remyelination promoting drugs. One limitation of snRNAseq is the lack of spatial resolution. This is of special interest for MS lesions, which may be spatially heterogeneous with respect to extent and composition of inflammation (such as mixed lesions) or remyelination (for example, partially remyelinated lesions). Spatial transcriptomics allows transcriptomic analyses in a spatially highly defined manner. Kaufmann et al. combined this technology with high sensitivity proteomics, published single cell-RNA sequencing data and in vivo perturbation model data to dissect underlying disease mechanisms of progressive MS and to identify new drug targets 49. They focussed their analysis on cortical grey matter from 13 progressive MS patients. There was a heterogenous distribution of neurodegeneration within the tissue and across patients, and pseudotemporal analyses allowed the identification of early neurodegenerative pathways in cortical MS tissue. Interestingly, these pathways were cell-type specific; however their regulation was highly connected between cell types suggesting multicellular pathological networks driving neurodegeneration. Kaufmann et al. confirmed earlier studies describing significant synaptic loss in cortical MS lesions and normal appearing grey matter (NAGM) 50. They also found an inverse correlation between genes regulating synaptic plasticity and maintenance and genes regulating inflammation and tissues remodelling expressed by astrocytes and innate immune cells. Interestingly, their data suggested that chronic inflammation only in combination with local failure of neurotrophic support result in full extent of multicomponent neurodegeneration. One caveat might be that the authors used pseudotemporal biostatistical approaches to identify early neurodegenerative events. It would be desirable to validate these results in tissue samples from MS patients with ongoing cortical demyelination; but these lesions are rarely available in post-mortem tissue collections. Using complex and elegant biostatistical analyses including in vivo CNS disease model data and their own spatial transcriptomic and proteomic results they identified and prioritized CNS enriched receptors as well as new pharmacological drug targets. (IV) Other demyelinating diseases Multiple sclerosis remains a diagnosis based on carefully considered clinical and MRI based criteria plus CSF findings of immunoglobulin (Ig) synthesis and oligoclonal bands. Although considered as an autoimmune disorder, no specific antigen-directed immune response is yet incorporated into the diagnostic criteria. However, over the last years, antigen specific immune responses have been identified in patients who would previously have been included within the MS umbrella. The first of these was NMO featuring AQP4directed antibodies. Consensus clinical syndromes and MRI findings related to optic nerve, spinal cord, area postrema, other brainstem, diencephalic, or cerebral presentations are now recognized 51. More recent has been recognition of monophasic or recurrent syndromes linked to presence of anti-MOG antibody. These expanding syndromes now include optic neuritis (ON), transverse myelitis (TM), encephalitis, and seizures, as well as aseptic meningitis and peripheral nervous system demyelination (reviewed in 52). MOGAD can be presented across the age spectrum. A specific question of interest addressed by Gaudioso et al. is the frequency of MOG-IgG and AQP4-IgG among patients with pediatric-onset multiple sclerosis (POMS) 53. In their review of 493 cases, MOG-IgG was positive in 30 cases (6%) and zero controls; none were NMO-antibody positive. Twenty-five of the 30 positive patients were judged to have MOGAD clinical criteria, 5 maintained a diagnosis of multiple sclerosis (MS). Only 44% had serum Epstein-Barr virus (EBV) positivity and only 20% had cerebrospinal fluid oligoclonal bands. These distinct “MS-related” disorders are receiving increasing attention with regard to optimal treatment paradigms, both for acute episodes and for prevention of recurrences. Cerebral cortical encephalitis (CCE) is a recently recognized phenotype of MOGAD. It is characterized by cortical T2-fluid-attenuated inversion recovery hyperintensity on MRI 54. Valencia-Sanchez et al. presented the clinical, imaging and pathological features of 19 CCE patients, identified by screening their cohort of 285 MOGAD patients 55. The majority of CCE occurred in patients with childhood onset of MOGAD and many patients presented with epilepsy. Pathology details were available in two patients. In one patient inactive perivascular demyelinated lesions in cortex and subcortical white matter were observed. The second biopsy was characterized by marked meningeal inflammation, extensive macrophage and microglia activation in the meninges as well as cortex and extensive subpial demyelination. The CD4/CD8 ratio was variable in these two cases and varied between 2.4 and 0.7. Few B cells aggregates were found in the meninges of one patient; terminal complement activation products were absent. Extensive subpial demyelination has been described to be specific for multiple sclerosis 56. This recent report, together with earlier MOGAD case reports demonstrate that subpial demyelination can also be found in other inflammatory demyelinating diseases and raises speculation regarding the role of humoral immune constituents and soluble inflammatory mediators as contributors 57,58. A further recently addressed issue is whether SARS-CoV-2 vaccination programs could result in increased rates of MOGAD. Francis et al reported a total of 25 cases referred to specialized MOGAD centers in the UK after the introduction of the COVID vaccination program; their conclusion was “these observations might support a causative role of the ChAdOx1S vaccine in inflammatory CNS disease and particularly MOGAD” 59. Summary The development of successful systemic immunomodulatory or anti-inflammatory treatment options for MS within the last two decades resulted in the discovery of progression independent of relapse activity. To dissect the underlying mechanisms driving this progression represents currently the biggest challenge in the field of MS. No animal model for the progressive disease phase of MS exists. Therefore, clinically well characterized human tissue collections and modern technologies, such as sc/snRNA sequencing and spatial transcriptomics will be essential to dissect the cellular and molecular mechanisms driving disease progression and to detect potential biomarkers. Clinical, imaging and other para-clinical studies are required to further validate candidate pathways and potential biomarkers. Here, we summarized some studies which started to disentangle the underlying pathways for disease progression and may serve as blueprint for future studies. References 1. Kappos, L. et al. Contribution of Relapse Independent Progression vs Relapse Associated Worsening to Overall Confirmed Disability Accumulation in Typical Relapsing Multiple Sclerosis in a Pooled Analysis of 2 Randomized Clinical Trials. JAMA Neurol 77, 1132 1140, https://doi.org/10.1001/jamaneurol.2020.1568 (2020). 2. University of California et al. Silent progression in disease activity free relapsing multiple sclerosis. Ann Neurol 85, 653 666, https://doi.org/10.1002/ana.25463 (2019). 3. Kuhlmann, T. et al. Multiple sclerosis progression: time for a new mechanism driven framework. Lancet Neurol 22, 78 88, https://doi.org/10.1016/S1474 4422(22)00289 7 (2023). 4. Macnair, W. et al. Single nuclei RNAseq stratifies multiple sclerosis patients into three distinct white matter glia responses. bioRxiv, https://doi.org/10.1101/2022.04.06.487263 (2022). 5. Tonietto, M. et al. Periventricular remyelination failure in multiple sclerosis: a substrate for neurodegeneration. Brain 146, 182 194, https://doi.org/10.1093/brain/awac334 (2023). 6. Bjornevik, K. et al. Longitudinal analysis reveals high prevalence of Epstein Barr virus associated with multiple sclerosis. Science 375, 296 301, https://doi.org/10.1126/science.abj8222 (2022). 7. Rostgaard, K., Nielsen, N. M., Melbye, M., Frisch, M. & Hjalgrim, H. Siblings reduce multiple sclerosis risk by preventing delayed primary Epstein Barr virus infection. Brain, https://doi.org/10.1093/brain/awac401 (2022). 8. Lanz, T. V. et al. Clonally expanded B cells in multiple sclerosis bind EBV EBNA1 and GlialCAM. Nature 603, 321 327, https://doi.org/10.1038/s41586 022 04432 7 (2022). 9. Hoftberger, R. & Lassmann, H. Immune mediated disorders. Handb Clin Neurol 145, 285 299, https://doi.org/10.1016/B978 0 12 802395 2.00020 1 (2017). 10. Wingerchuk, D. M. & Lucchinetti, C. F. Neuromyelitis Optica Spectrum Disorder. N Engl J Med 387, 631 639, https://doi.org/10.1056/NEJMra1904655 (2022). 11. Schneider Hohendorf, T. et al. Broader Epstein Barr virus specific T cell receptor repertoire in patients with multiple sclerosis. J Exp Med 219, https://doi.org/10.1084/jem.20220650 (2022). 12. Ingelfinger, F. et al. Twin study reveals non heritable immune perturbations in multiple sclerosis. Nature 603, 152 158, https://doi.org/10.1038/s41586 022 04419 4 (2022). 13. iMSM Consortium. Gut microbiome of multiple sclerosis patients and paired household healthy controls reveal associations with disease risk and course. Cell 185, 3467 3486 e3416, https://doi.org/10.1016/j.cell.2022.08.021 (2022). 14. Hosang, L. et al. The lung microbiome regulates brain autoimmunity. Nature 603, 138 144, https://doi.org/10.1038/s41586 022 04427 4 (2022). 15. Dorion, M. F. et al. Systematic comparison of culture media uncovers phenotypic shift of primary human microglia defined by reduced reliance to CSF1R signaling. Glia, https://doi.org/10.1002/glia.24338 (2023). 16. Thompson, A. J. et al. Diagnosis of multiple sclerosis: 2017 revisions of the McDonald criteria. Lancet Neurol 17, 162 173, https://doi.org/10.1016/S1474 4422(17)30470 2 (2018). 17. Lublin, F. D. et al. Defining the clinical course of multiple sclerosis: the 2013 revisions. Neurology 83, 278 286, https://doi.org/10.1212/WNL.0000000000000560 (2014). 18. Lublin, F. D. et al. How patients with multiple sclerosis acquire disability. Brain 145, 3147 3161, https://doi.org/10.1093/brain/awac016 (2022). 19. Tur, C. et al. Association of Early Progression Independent of Relapse Activity With Long term Disability After a First Demyelinating Event in Multiple Sclerosis. JAMA Neurol, https://doi.org/10.1001/jamaneurol.2022.4655 (2022). 20. Beynon, V. et al. Chronic lesion activity and disability progression in secondary progressive multiple sclerosis. BMJ Neurol Open 4, e000240, https://doi.org/10.1136/bmjno 2021 000240 (2022). 21. Bevan, R. J. et al. Meningeal inflammation and cortical demyelination in acute multiple sclerosis. Ann Neurol 84, 829 842, https://doi.org/10.1002/ana.25365 (2018). 22. Magliozzi, R. et al. A Gradient of neuronal loss and meningeal inflammation in multiple sclerosis. Ann. Neurol 68, 477 493, https://doi.org/10.1002/ana.22230 (2010). 23. Magliozzi, R. et al. Meningeal B cell follicles in secondary progressive multiple sclerosis associate with early onset of disease and severe cortical pathology. Brain 130, 1089 1104, https://doi.org/10.1093/brain/awm038 (2007). 24. Howell, O. W. et al. Meningeal inflammation is widespread and linked to cortical pathology in multiple sclerosis. Brain 134, 2755 2771, https://doi.org/10.1093/brain/awr182 (2011). 25. Ahmed, S. M. et al. Accumulation of meningeal lymphocytes correlates with white matter lesion activity in progressive multiple sclerosis. JCI Insight 7, https://doi.org/10.1172/jci.insight.151683 (2022). 26. Howell, O. W. et al. Extensive grey matter pathology in the cerebellum in multiple sclerosis is linked to inflammation in the subarachnoid space. Neuropathol Appl Neurobiol 41, 798 813, https://doi.org/10.1111/nan.12199 (2015). 27. Fransen, N. L. et al. Absence of B Cells in Brainstem and White Matter Lesions Associates With Less Severe Disease and Absence of Oligoclonal Bands in MS. Neurol Neuroimmunol Neuroinflamm 8, https://doi.org/10.1212/NXI.0000000000000955 (2021). 28. Reali, C. et al. B cell rich meningeal inflammation associates with increased spinal cord pathology in multiple sclerosis. Brain Pathol 30, 779 793, https://doi.org/10.1111/bpa.12841 (2020). 29. Drieu, A. et al. Parenchymal border macrophages regulate the flow dynamics of the cerebrospinal fluid. Nature 611, 585 593, https://doi.org/10.1038/s41586 022 05397 3 (2022). 30. Carotenuto, A. et al. Glymphatic system impairment in multiple sclerosis: relation with brain damage and disability. Brain 145, 2785 2795, https://doi.org/10.1093/brain/awab454 (2022). 31. Fabriek, B. O. et al. In vivo detection of myelin proteins in cervical lymph nodes of MS patients using ultrasound guided fine needle aspiration cytology. J Neuroimmunol 161, 190 194, https://doi.org/10.1016/j.jneuroim.2004.12.018 (2005). 32. Merlini, A. et al. Distinct roles of the meningeal layers in CNS autoimmunity. Nat Neurosci 25, 887 899, https://doi.org/10.1038/s41593 022 01108 3 (2022). 33. van den Bosch, A. M. R. et al. Ultrastructural Axon Myelin Unit Alterations in Multiple Sclerosis Correlate with Inflammation. Ann Neurol, https://doi.org/10.1002/ana.26585 (2022). 34. Wolswijk, G. & Balesar, R. Changes in the expression and localization of the paranodal protein Caspr on axons in chronic multiple sclerosis. Brain 126, 1638 1649, https://doi.org/10.1093/brain/awg151 (2003). 35. Gallego Delgado, P. et al. Neuroinflammation in the normal appearing white matter (NAWM) of the multiple sclerosis brain causes abnormalities at the nodes of Ranvier. PLoS Biol 18, e3001008, https://doi.org/10.1371/journal.pbio.3001008 (2020). 36. Irvine, K. A. & Blakemore, W. F. Remyelination protects axons from demyelination associated axon degeneration. Brain 131, 1464 1477, https://doi.org/10.1093/brain/awn080 (2008). 37. Duncan, I. D., Brower, A., Kondo, Y., Curlee, J. F., Jr. & Schultz, R. D. Extensive remyelination of the CNS leads to functional recovery. Proc. Natl. Acad. Sci. U. S. A 106, 6832 6836, https://doi.org/10.1073/pnas.0812500106 (2009). 38. Sarrazin, N. et al. Failed remyelination of the nonhuman primate optic nerve leads to axon degeneration, retinal damages, and visual dysfunction. Proc Natl Acad Sci U S A 119, e2115973119, https://doi.org/10.1073/pnas.2115973119 (2022). 39. Patrikios, P. et al. Remyelination is extensive in a subset of multiple sclerosis patients. Brain 129, 3165 3172, https://doi.org/10.1093/brain/awl217 (2006). 40. Goldschmidt, T., Antel, J., Konig, F. B., Brück, W. & Kuhlmann, T. Remyelination capacity of the MS brain decreases with disease chronicity. Neurology 72, 1914 1921, https://doi.org/10.1212/WNL.0b013e3181a8260a (2009). 41. Arnold, D. L. et al. Effect of siponimod on magnetic resonance imaging measures of neurodegeneration and myelination in secondary progressive multiple sclerosis: Gray matter atrophy and magnetization transfer ratio analyses from the EXPAND phase 3 trial. Mult Scler 28, 1526 1540, https://doi.org/10.1177/13524585221076717 (2022). 42. Rahmanzadeh, R. et al. A New Advanced MRI Biomarker for Remyelinated Lesions in Multiple Sclerosis. Ann Neurol 92, 486 502, https://doi.org/10.1002/ana.26441 (2022). 43. Absinta, M. et al. Persistent 7 tesla phase rim predicts poor outcome in new multiple sclerosis patient lesions. J Clin Invest 126, 2597 2609, https://doi.org/10.1172/JCI86198 (2016). 44. Chen, W. et al. Quantitative susceptibility mapping of multiple sclerosis lesions at various ages. Radiology 271, 183 192, https://doi.org/10.1148/radiol.13130353 (2014). 45. Chen, J. T., Collins, D. L., Atkins, H. L., Freedman, M. S. & Arnold, D. L. Magnetization transfer ratio evolution with demyelination and remyelination in multiple sclerosis lesions. Ann. Neurol 63, 254 262, https://doi.org/10.1002/ana.21302 (2008). 46. Jakel, S. et al. Altered human oligodendrocyte heterogeneity in multiple sclerosis. Nature 566, 543 547, https://doi.org/10.1038/s41586 019 0903 2 (2019). 47. Schirmer, L. et al. Neuronal vulnerability and multilineage diversity in multiple sclerosis. Nature 573, 75 82, https://doi.org/10.1038/s41586 019 1404 z (2019). 48. Absinta, M. et al. A lymphocyte microglia astrocyte axis in chronic active multiple sclerosis. Nature 597, 709 714, https://doi.org/10.1038/s41586 021 03892 7 (2021). 49. Kaufmann, M. et al. Identification of early neurodegenerative pathways in progressive multiple sclerosis. Nat Neurosci 25, 944 955, https://doi.org/10.1038/s41593 022 01097 3 (2022). 50. Jurgens, T. et al. Reconstruction of single cortical projection neurons reveals primary spine loss in multiple sclerosis. Brain 139, 39 46, https://doi.org/10.1093/brain/awv353 (2016). 51. Wingerchuk, D. M. et al. International consensus diagnostic criteria for neuromyelitis optica spectrum disorders. Neurology 85, 177 189,https://doi.org/10.1212/WNL.0000000000001729 (2015). 52. Marignier, R. et al. Myelin oligodendrocyte glycoprotein antibody associated disease. Lancet Neurol 20, 762 772, https://doi.org/10.1016/S1474 4422(21)00218 0 (2021). 53. Gaudioso, C. M. et al. MOG and AQP4 Antibodies among Children with Multiple Sclerosis and Controls. Ann Neurol, https://doi.org/10.1002/ana.26502 (2022). 54. Ogawa, R. et al. MOG antibody positive, benign, unilateral, cerebral cortical encephalitis with epilepsy. Neurol Neuroimmunol Neuroinflamm 4, e322, https://doi.org/10.1212/NXI.0000000000000322 (2017). 55. Valencia Sanchez, C. et al. Cerebral Cortical Encephalitis in Myelin Oligodendrocyte Glycoprotein Antibody Associated Disease. Ann Neurol, https://doi.org/10.1002/ana.26549 (2022). 56. Junker, A. et al. Extensive subpial cortical demyelination is specific to multiple sclerosis. Brain Pathol 30, 641 652, https://doi.org/10.1111/bpa.12813 (2020). 57. Hoftberger, R. et al. The pathology of central nervous system inflammatory demyelinating disease accompanying myelin oligodendrocyte glycoprotein autoantibody. Acta Neuropathol 139, 875 892, https://doi.org/10.1007/s00401 020 02132 y (2020). 58. Takai, Y. et al. Myelin oligodendrocyte glycoprotein antibody associated disease: an immunopathological study. Brain 143, 1431 1446, https://doi.org/10.1093/brain/awaa102 (2020). 59. Francis, A. G. et al. Acute Inflammatory Diseases of the Central Nervous System After SARS CoV 2 Vaccination. Neurol Neuroimmunol Neuroinflamm 10, https://doi.org/10.1212/NXI.0000000000200063 (2023).
Copyright: © 2023 The author(s). This is an open access article distributed under the terms of the Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited, a link to the Creative Commons license is provided, and any changes are indicated. The Creative Commons Public Domain Dedication waiver (https://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated. |