|
|
|
Free Neuropathology 2:18 (2021) |
|
Opinion Piece |
|
SARS-CoV-2 vaccination induced cerebral venous sinus thrombosis: Do megakaryocytes, platelets and lipid mediators make up the orchestra? |
|
Kate Chander Chiang1, Ravi Raghavan2, Ajay Gupta1,3 |
|
1 KARE Biosciences, Orange, CA 92869, USA |
|
Corresponding author: |
|
Submitted: 6 June 2021 Accepted: 1 July 2021 Copyedited by: Biswarathan Ramani Published: 7 July 2021 |
|
Keywords: SARS-CoV-2, COVID-19, Vaccine, Thrombosis, Cerebral venous sinus thrombosis, Aspirin, Thromboxane, COX-2, Adenoviral vector, Megakaryocyte |
|
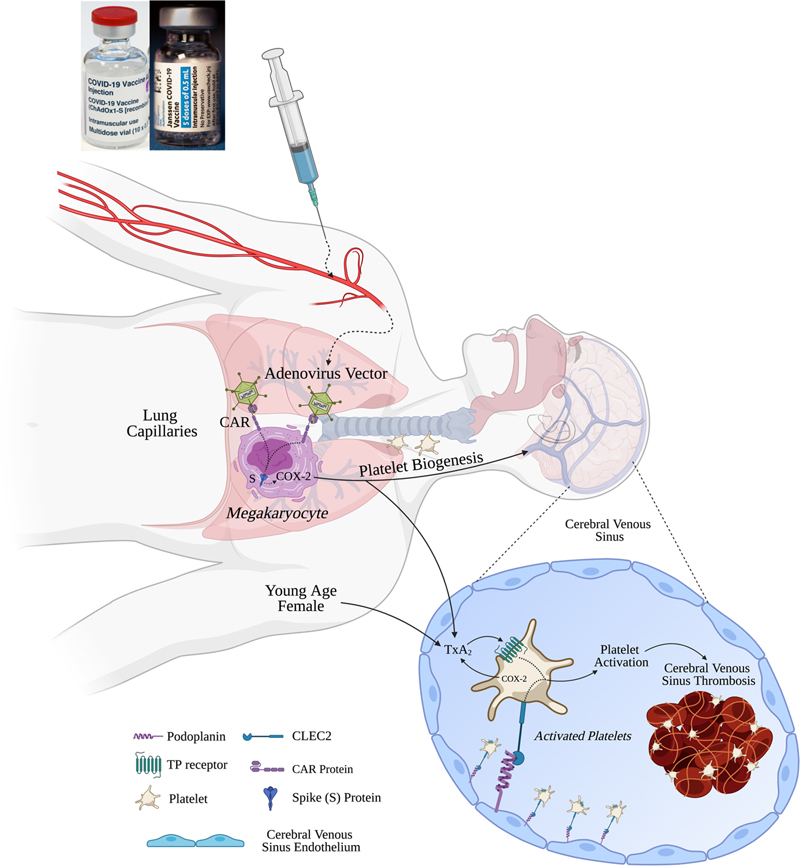
Abstract The COVID-19 vaccines comprised of adenoviral vectors encoding the Spike (S) glycoprotein of SARS-CoV-2 are highly effective but associated with rare thrombotic complications. The adenovirus vector infects epithelial cells expressing the coxsackievirus and adenovirus receptor (CAR). The S glycoprotein expressed locally stimulates neutralizing antibody and cellular immune responses. These vaccines have been associated with thromboembolic events including cerebral venous sinus thrombosis (CVST). S glycoprotein stimulates the expression of cyclooxygenase-2 (COX-2) and leads to massive generation of thromboxane A2 in COVID-19. Megakaryocytes express CAR and we postulate that S glycoprotein stimulated generation of thromboxane A2 leads to megakaryocyte activation, biogenesis of activated platelets and thereby increased thrombogenicity. Cerebral vein sinuses express podoplanin, a natural ligand for CLEC2 receptors on platelets. Platelets traversing through the cerebral vein sinuses could be further activated by thromboxane A2-dependent podoplanin-CLEC2 signaling, leading to CVST. A prothrombotic hormonal milieu, and increased generation of thromboxane A2 and platelet activation in healthy females compared to males is consistent with increased risk for CVST observed in women. We propose that antiplatelet agents targeting thromboxane A2 receptor signaling such as low-dose aspirin merit consideration for chemoprophylaxis when administering the adenovirus based COVID-19 vaccines to young adults at risk of thrombosis provided there are no contraindications. Abbreviations TxA2 - thromboxane A2, DIC - disseminated intravascular coagulopathy, COX - cyclooxygenase, TTP - thrombotic thrombocytopenic purpura, CVST - cerebral venous sinus thrombosis, CLEC - C-type lectin-like receptor - TLR, toll-like receptor, CAR - coxsackievirus and adenovirus receptor, COVID-19 - coronavirus disease 2019, SARS-CoV-2 - severe acute respiratory syndrome coronavirus 2, PF4 - platelet factor 4 Introduction COVID-19 disease is caused by a novel positive-strand RNA coronavirus (SARS-CoV-2), which belongs to the Coronaviridae family, along with the severe acute respiratory syndrome (SARS) and the Middle East respiratory syndrome (MERS) coronaviruses.1 The genome of these viruses encodes several non-structural and structural proteins, including spike, envelope, membrane, and nucleocapsid proteins.2 The majority of the vaccines for COVID-19 that employ administration of viral antigens or viral gene sequences aim to induce neutralizing antibodies against the viral spike protein (S), preventing uptake through the ACE2 receptor, and thereby blocking infection.3 The Janssen COVID-19 vaccine (Johnson & Johnson) is comprised of a recombinant, replication- incompetent Ad26 vector, encoding a stabilized variant of the SARS-CoV-2 spike protein. The ChAdOx1 nCoV-19 vaccine (AZD1222, Vaxzevria®) was developed at Oxford University and consists of a replication-deficient chimpanzee adenoviral vector ChAdOx1, encoding the S protein.4 In US Phase III trials, Vaxzevria has been demonstrated to have 79% efficacy at preventing symptomatic COVID-19, and 100% efficacy against severe or critical disease and hospitalization, with comparable efficacy across ethnicity, gender and age.5 However, Vaxzevria has been associated with thrombotic and embolic events including disseminated intravascular coagulation (DIC) and cerebral venous sinus thrombosis (CVST), occurring within 14 days after vaccination, mostly in people under 55 years of age, the majority of whom have been women.6 Data from Europe suggests that the event rate for thromboembolic events may be about 10 per million vaccinated. Antibodies to platelet factor 4 (PF4)/heparin complexes have been recently reported in a few patients.7 However, the significance of this finding remains to be established. As of April 12, 2021, about 6.8 million doses of the Janssen vaccine have been administered in the U.S.8 CDC and FDA are reviewing data involving six reported U.S. cases of CVST in combination with thrombocytopenia.8 All six cases occurred among women between the ages of 18 and 48, and symptoms occurred 6 to 13 days after vaccination.8 SARS-CoV-2 is known to cause thromboinflammation leading to thrombotic microangiopathy, pulmonary thrombosis, pedal acro-ischemia (“COVID-toes”), arterial clots, strokes, cardiomyopathy, coronary and systemic vasculitis, deep venous thrombosis, pulmonary embolism, and microvascular thrombosis in renal, cardiac and brain vascularture.9-14 Cerebral venous sinus thrombosis (CVST) has also been reported in COVID-19 patients.15 Amongst 34,331 hospitalized COVID-19 patients, CVST was diagnosed in 28.16 In a multicenter, multinational, cross sectional, retrospective study of 8 patients diagnosed with CVST and COVID-19, seven were women with an average age of 63 years.17 In a combined series of 41 patients with COVID-19 and CVST, the average age was about 50 years (SD, 16.5 years).17 The pathobiology of thrombotic events associated with the AstraZeneca vaccine should be viewed in the context of mechanisms underlying thromboinflammation that complicates SARS-CoV-2 infection and COVID-19 disease. A. Role of COX-2 and thromboxane A2 in thromboinflammation complicating adenovirus-based COVID-19 vaccine encoding the Spike protein of SARS-CoV-2 Thromboinflammation in COVID-19 seems to be primarily caused by endothelial, platelet and neutrophil activation, platelet-neutrophil aggregates and release of neutrophil extracellular traps (NETs).13,18 Platelet activation in COVID-19 is fueled by a lipid storm characterized by massive increases in thromboxane A2 (TxA2) levels in the blood and bronchoalveolar lavage fluid.19,20 Cyclooxygenase (COX) enzymes catalyze the first step in the biosynthesis of thromboxane A2 from arachidonic acid, and COX-2 expression is induced by the spike protein of coronaviruses.21 We postulate that an aberrant increase in thromboxane A2 generation induced by the spike protein expression from adenovirus vaccines leads to thromboinflammation and CVST. The support for the above proposed mechanism comes from the following observations. First, when mice of different age groups were infected with SARS-CoV virus, the generation of TxA2 was markedly increased in younger mice compared to middle aged mice.22 Furthermore, in children with asymptomatic or mildly symptomatic SARS-CoV-2 infection, microvascular thrombosis and thrombotic microangiopathy occur early in infection.19 These observations are consistent with the higher risk for thrombosis in adults under 60 years of age, compared with the older age group.6,7 Second, platelets from female mice are much more reactive than from male mice.23 Furthermore, TxA2 generation, TxA2-platelet interaction and platelet activation is increased in women compared to men.24,25 These observations are consistent with disproportionately increased risk of thrombosis in women following AstraZeneca and Janssen COVID-19 vaccines. The adenoviral vector ChAdOx1, containing the nCoV-19 spike protein gene, infects host cells through the coxsackievirus and adenovirus receptor (CAR).26 CAR-dependent cell entry of the viral vector allows insertion of the SARS-CoV-2 spike protein gene and expression of spike protein by host cells (Figure 1). CAR is primarily expressed on epithelial tight junctions including on enterocytes.27,28 CAR expression has also been reported in platelets,29 and, since platelets are anucleate cells, CAR expression by megakaryocytes can be inferred. Therefore, AstraZeneca and Janssen vaccines would be expected to induce expression of spike protein in megakaryocytes, platelets (Figure 1) and enterocytes. We postulate that the expression of CAR receptors by basolateral membrane on enterocytes allows uptake of adenovirus vector,28 leading to spike protein-induced COX-2 expression and generation of thromboxane A2. Delivery of thromboxane A2 into the splanchnic circulation could activate platelets locally, leading to splanchnic vein thrombosis associated with adenovirus vaccines.30
Figure 1. AstraZeneca or Janssen COVID-19 vaccine-induced thromboinflammation and cerebral venous sinus thrombosis (CVST) - Proposed Mechanisms: Adenovirus carrier delivers SARS-CoV-2 DNA encoding the spike (S) protein to the lung megakaryocytes via the coxsackie-adenovirus receptor (CAR). Spike protein induces COX-2 expression in megakaryocytes leading to megakaryocyte activation, biogenesis of activated platelets that express COX-2 and generate thromboxane A2 (TxA2). Cerebral venous sinuses express podoplanin, a natural ligand for CLEC2 receptors on platelets. Platelets traversing through the cerebral venous sinuses would be further activated by thromboxane A2-dependent podoplanin-CLEC2 signaling, leading to release of extracellular vesicles, thereby promoting CLEC5A- and TLR2-mediated neutrophil activation, thromboinflammation and CVST. Young age and female gender are associated with increased TxA2 generation and platelet activation, respectively, and hence increased risk of thromboembolic complications following SARS-CoV-2 vaccination with adenovirus-based vaccines. Spike protein of coronaviruses is known to induce COX-2 gene expression.21,31 COX-2 expression is induced during normal human megakaryopoiesis and characterizes newly formed platelets.32 While in healthy controls <10% of circulating platelets express COX-2, in patients with high platelet generation, up to 60% of platelets express COX-2.32 Generation of TxA2 by platelets is markedly suppressed by COX-2 inhibition in patients with increased megakaryopoiesis versus healthy subjects.32 Therefore, we postulate that expression of spike protein induces COX-2 expression and generation of thromboxane A2 by megakaryocytes. TxA2 promotes biogenesis of activated platelets expressing COX-2. Platelet TxA2 generation leads to platelet activation and aggregation, and thereby thromboinflammation (Figure 1). Extravascular spaces of the lungs comprise populations of mature and immature megakaryocytes that originate from the bone marrow, such that lungs are a major site of platelet biogenesis, accounting for approximately 50% of total platelet production or about 10 million platelets per hour.33 More than 1 million extravascular megakaryocytes have been observed in each lung of transplant mice.33 Following intramuscular injection of the AstraZeneca and Janssen vaccines, the adenovirus vector will traverse the veins and lymphatics to be delivered to the pulmonary circulation, thereby exposing lung megakaryocytes in the first pass. Interestingly, under thrombocytopenic conditions, haematopoietic progenitors migrate out of the lung to repopulate the bone marrow and completely reconstitute blood platelet count.33 Since thrombocytopenia is a common feature of CVST associated with adenovirus SARS-CoV-2 vaccines, migration of megakaryocytes from the lungs into the systemic circulation including the cerebral circulation could further predispose to CVST. In this regard, it is relevant that megakaryocytes have been found in the cerebral circulation in patients deceased from severe COVID-19.34 The prothrombotic mechanisms postulated above could be operative alone or in conjunction with other underlying factors that have been proposed including the presence of anti-platelet factor 4 (PF4) antibodies as reported by Greinacher and others.7 Notably, amongst 31 cases of CVST in close temporal relationship with a COVID-19 vaccination, anti-PF4 antibodies were positive in 22 cases and negative in 9.35 Therefore, CVST following COVID-19 vaccination appears to be a multifactorial disease. Further studies using cultured megakaryocytes and animal models are needed to define the underlying mechanisms. Even though the risk of CVST is increased about 10-fold by the adenovirus based COVID-19 vaccines,36 it remains unclear why CVST remains a rare event. To further define the importance of these causative factors, biomarkers such as plasma thromboxane B2, anti-PF4 antibodies and anti-phospholipid antibodies should be measured in the thrombosis-free vaccinated population. Such studies may help define the role of anti-PF4 antibodies as causative versus an epiphenomenon. Interestingly, almost all patients with thrombosis and thrombocytopenia were found to have anti-PF4 antibodies.35 On the other hand, majority of patients with thrombosis without thrombocytopenia lacked anti-PF4 antibodies.35 The postulated mechanisms may be especially relevant to thrombotic events occurring in the absence of thrombocytopenia or anti-PF4 antibodies. B. Predilection of cerebral venous sinuses for thrombosis following vaccination Recent studies have demonstrated that arterial, venous and sinusoidal endothelial cells in the brain uniquely express markers of the lymphatic endothelium including podoplanin.37 Podoplanin (D2-40) serves as a ligand for CLEC2 receptors on platelets.38 Thromboxane A2-dependent CLEC2 signaling leads to platelet activation (Figure 1), while a TxA2 receptor antagonist nearly abolished CLEC2 signaling and platelet activation.38 TxA2-dependent CLEC2 signaling promotes release of exosomes and microvesicles from platelets, leading to activation of CLEC5A and TLR2 receptors respectively on neutrophils, neutrophil activation and release of neutrophil extracellular traps (NETs) (Figure 1).39 Neutrophil activation, more than platelet activation, is associated with thrombotic complications in COVID-19.13,18,40 As proposed above, the expression of podoplanin by cerebral venous sinuses may be responsible for the predilection of brain vascular bed to thromboinflammation and CVST as a complication of COVID-19 vaccines. The valves at the lymphovenous junction and the podoplanin-CLEC2 lymphovenous hemostasis act as “fail-safe” mechanisms to prevent movement of blood into the lymphatic system, thereby preventing coagulation in the lymphatics despite expression of podoplanin in the lymphatic endothelium.41 C. Chemoprophylaxis with antiplatelet agents In animal models of endotoxin-mediated endothelial injury and thromboinflammation, antagonism of thromboxane A2 signaling prevents ARDS, reduces myocardial damage and increases survival.42-44 Considering the key role played by platelets in thromboinflammation, we propose consideration of anti-platelet agents, either aspirin or TxA2 receptor antagonists, as chemoprophylactic agents when the AstraZeneca vaccine is administered to adults between 18 and 60 years of age.45 High bleeding risk because of another medical condition or medication would be contraindications to use of antiplatelet agents.45 Medical conditions that increase bleeding risk include previous gastrointestinal bleeding, peptic ulcer disease, blood clotting problems, and kidney disease.45 Medications that increase bleeding risk include nonsteroidal anti-inflammatory drugs, steroids, and other anticoagulants or anti-platelet agents.45 Aspirin appears to be safe in COVID-19. In a retrospective observational study in hospitalized patients with COVID-19, low-dose aspirin was found to be effective in reducing morbidity and mortality and was not associated with any safety issues including major bleeding.46 Therefore, aspirin is likely to be safe as an adjunct to COVID-19 vaccines even in the event of a subsequent infection with SARS-CoV-2 virus. Can aspirin influence the host immune response to the COVID-19 vaccines? This issue merits further investigation. When healthy adults > 65 years of age were given influenza vaccine and randomized to receive 300 mg aspirin or placebo on days 1, 2, 3, 5 and 7, the aspirin group showed 4-fold or greater rise in influenza specific antibodies.47 The risk-benefit analysis, based on above information, suggests that a one to three week course of low-dose aspirin merits consideration in order to prevent the thromboembolic events associated with the AstraZeneca vaccine. Summary Thromboembolic disease, CVST and thrombocytopenia have been reported in association with AstraZeneca and Janssen COVID-19 vaccines, especially in younger women. Many countries have halted use of these vaccines for adults under 30 to 60 years of age. European and North American countries generally have access to mRNA vaccines. However, in many Asian and African countries the choices are limited to adenovirus-based COVID-19 vaccines. The governments in such countries are forging ahead with vaccinating all adults, including those under 60 years of age, with Vaxzevria, Covishield (the version of Vaxzevria manufactured by the Serum Institute of India) or the Janssen vaccines. This has led to grave concern and anxiety amongst the citizens and medical professionals. Considering the profound global public health implications of limiting the use of these vaccines, it is critical to understand the pathobiology of vaccination induced thrombotic events in order to guide strategies aimed at prevention. In this regard, studies are urgently needed to examine lipid mediators and the thromboxane A2 - platelet axis following vaccination with these vaccines compared with mRNA vaccines. The risk-benefit analysis based on information presented here suggests that chemoprophylaxis using a short course of low-dose aspirin in adults under 60 years of age and lacking any contraindications to aspirin may be justified in conjunction with adenovirus-based COVID-19 vaccines in order to prevent thromboembolic events and enhance safety. Acknowledgment We thank Prof. Andrzej Breborowicz, MD, PhD, Department of Pathophysiology, Poznan University of Medical Sciences, Poland for a careful review and critique of this work. Funding No funding was required. Conflict of Interest AG and KCC have filed a patent for use of Ramatroban as an anti-thrombotic and immune modulator in SARS-CoV-2 infection. The patents have been licensed to KARE Biosciences. KCC is an employee of KARE Biosciences. Author Contributions AG and KCC conceptualized, created the framework and drafted the original version. All authors reviewed and edited the final version. References 1. Ortiz-Prado E, Simbaña-Rivera K, Gómez-Barreno L, et al. Clinical, molecular, and epidemiological characterization of the SARS-CoV-2 virus and the Coronavirus Disease 2019 (COVID-19), a comprehensive literature review. Diagnostic Microbiology and Infectious Disease. 2020;98(1):115094. 2. Du L, He Y, Zhou Y, Liu S, Zheng B-J, Jiang S. The spike protein of SARS-CoV — a target for vaccine and therapeutic development. Nature Reviews Microbiology. 2009;7(3):226-236. 3. Kyriakidis NC, López-Cortés A, González EV, Grimaldos AB, Prado EO. SARS-CoV-2 vaccines strategies: a comprehensive review of phase 3 candidates. npj Vaccines. 2021;6(1). 4. Voysey M, Clemens SAC, Madhi SA, et al. Safety and efficacy of the ChAdOx1 nCoV-19 vaccine (AZD1222) against SARS-CoV-2: an interim analysis of four randomised controlled trials in Brazil, South Africa, and the UK. The Lancet. 2021;397(10269):99-111. 5. AstraZeneca. AZD1222 US Phase III trial met primary efficacy endpoint in preventing COVID-19 at interim analysis. https://www.astrazeneca.com/media-centre/press-releases/2021/astrazeneca-us-vaccine-trial-met-primary-endpoint.html. Published 2021. Accessed April 5, 2021. 6. European Medicines Agency. COVID-19 vaccine safety update VAXZEVRIA. https://www.ema.europa.eu/en/documents/covid-19-vaccine-safety-update/covid-19-vaccine-safety-update-vaxzevria-previously-covid-19-vaccine-astrazeneca-29-march-2021_en.pdf. Published 2021. Accessed April 4, 2021. 7. Greinacher A, Thiele T, Warkentin TE, Weisser K, Kyrle PA, Eichinger S. Thrombotic Thrombocytopenia after ChAdOx1 nCov-19 Vaccination. New England Journal of Medicine. 2021;384(22):2092-2101. 8. CDC. Joint CDC and FDA Statement on Johnson & Johnson COVID-19 Vaccine. https://www.cdc.gov/media/releases/2021/s0413-JJ-vaccine.html. Published 2021. Accessed April 13, 2021. 9. Ackermann M, Verleden SE, Kuehnel M, et al. Pulmonary Vascular Endothelialitis, Thrombosis, and Angiogenesis in Covid-19. New England Journal of Medicine. 2020;383(2):120-128. 10. Goyal P, Choi JJ, Pinheiro LC, et al. Clinical Characteristics of Covid-19 in New York City. New England Journal of Medicine. 2020;382(24):2372-2374. 11. Guan W-J, Ni Z-Y, Hu Y, et al. Clinical Characteristics of Coronavirus Disease 2019 in China. New England Journal of Medicine. 2020;382(18):1708-1720. 12. Hottz ED, Azevedo-Quintanilha IG, Palhinha L, et al. Platelet activation and platelet-monocyte aggregates formation trigger tissue factor expression in severe COVID-19 patients. Blood. 2020;136(11):1330-1341. 13. Nicolai L, Leunig A, Brambs S, et al. Immunothrombotic Dysregulation in COVID-19 Pneumonia is Associated with Respiratory Failure and Coagulopathy. Circulation. 2020;142(12):1176-1189. 14. Song W-C, Fitzgerald GA. COVID-19, microangiopathy, hemostatic activation, and complement. Journal of Clinical Investigation. 2020;130(8):3950-3953. 15. Mowla A, Shakibajahromi B, Shahjouei S, et al. Cerebral venous sinus thrombosis associated with SARS-CoV-2; a multinational case series. Journal of Neurological Sciences. 2020;419:117183. 16. Baldini T, Asioli GM, Romoli M, et al. Cerebral venous thrombosis and severe acute respiratory syndrome coronavirus-2 infection: A systematic review and meta-analysis. European Journal of Neurology. 2021;10.1111/ene.14727. 17. Abdalkader M, Shaikh SP, Siegler JE, et al. Cerebral Venous Sinus Thrombosis in COVID-19 Patients: A Multicenter Study and Review of Literature. Journal of Stroke and Cerebrovascular Diseases. 2021;30(6):105733. 18. Petito E, Falcinelli E, Paliani U, et al. Association of Neutrophil Activation, More Than Platelet Activation, With Thrombotic Complications in Coronavirus Disease 2019. The Journal of Infectious Diseases. 2021;223(6):933-944. 19. Diorio C, McNerney KO, Lambert M, et al. Evidence of thrombotic microangiopathy in children with SARS-CoV-2 across the spectrum of clinical presentations. Blood Advances. 2020;4(23):6051-6063. 20. Archambault AS, Zaid Y, Rakotoarivelo V, et al. High levels of eicosanoids and docosanoids in the lungs of intubated COVID‐19 patients. The FASEB Journal. 2021;35(6). 21. Liu M, Gu C, Wu J, Zhu Y. Amino acids 1 to 422 of the spike protein of SARS associated coronavirus are required for induction of cyclooxygenase-2. Virus Genes. 2006;33(3):309-317. 22. Vijay R, Hua X, Meyerholz DK, et al. Critical role of phospholipase A2 group IID in age-related susceptibility to severe acute respiratory syndrome-CoV infection. Journal of Experimental Medicine. 2015;212(11):1851-1868. 23. Leng X-H, Hong SY, Larrucea S, et al. Platelets of Female Mice Are Intrinsically More Sensitive to Agonists Than Are Platelets of Males. Arteriosclerosis, Thrombosis, and Vascular Biology. 2004;24(2):376-381. 24. Kim BS, Auerbach DA, Sadhra H, et al. A Sex-Specific Switch in Platelet Receptor Signaling Following Myocardial Infarction. In: Cold Spring Harbor Laboratory; 2019. 25. Eikelboom JW, Hirsh J, Weitz JI, Johnston M, Yi Q, Yusuf S. Aspirin-resistant thromboxane biosynthesis and the risk of myocardial infarction, stroke, or cardiovascular death in patients at high risk for cardiovascular events. Circulation. 2002;105(14):1650-1655. 26. Cohen CJ, Xiang ZQ, Gao G-P, Ertl HCJ, Wilson JM, Bergelson JM. Chimpanzee adenovirus CV-68 adapted as a gene delivery vector interacts with the coxsackievirus and adenovirus receptor. Journal of General Virology. 2002;83(1):151-155. 27. Cohen CJ, Shieh JT, Pickles RJ, Okegawa T, Hsieh JT, Bergelson JM. The coxsackievirus and adenovirus receptor is a transmembrane component of the tight junction. Proceedings of the National Academy of Sciences. 2001;98(26):15191-15196. 28. Kesisoglou F, Schmiedlin-Ren P, Fleisher D, Zimmermann EM. Adenoviral Transduction of Enterocytes and M-Cells Using in Vitro Models Based on Caco-2 Cells: The Coxsackievirus and Adenovirus Receptor (CAR) Mediates Both Apical and Basolateral Transduction. Molecular Pharmaceutics. 2010;7(3):619-629. 29. Assinger A. Platelets and infection - an emerging role of platelets in viral infection. Frontiers in Immunology. 2014;5:649. 30. Porres-Aguilar M, Lazo-Langner A, Panduro A, Uribe M. COVID-19 vaccine-induced immune thrombotic thrombocytopenia: An emerging cause of splanchnic vein thrombosis. Annals of Hepatology. 2021;23:100356. 31. Yan X, Hao Q, Mu Y, et al. Nucleocapsid protein of SARS-CoV activates the expression of cyclooxygenase-2 by binding directly to regulatory elements for nuclear factor-kappa B and CCAAT/enhancer binding protein. The International Journal of Biochemistry & Cell Biology. 2006;38(8):1417-1428. 32. Rocca B, Secchiero P, Ciabattoni G, et al. Cyclooxygenase-2 expression is induced during human megakaryopoiesis and characterizes newly formed platelets. Proceedings of the National Academy of Sciences. 2002;99(11):7634-7639. 33. Lefrançais E, Ortiz-Muñoz G, Caudrillier A, et al. The lung is a site of platelet biogenesis and a reservoir for haematopoietic progenitors. Nature. 2017;544(7648):105-109. 34. Nauen DW, Hooper JE, Stewart CM, Solomon IH. Assessing Brain Capillaries in Coronavirus Disease 2019. JAMA Neurology. 2021;78(6):760-762 35. Schultz NH, Sørvoll IH, Michelsen AE, et al. Thrombosis and Thrombocytopenia after ChAdOx1 nCoV-19 Vaccination. New England Journal of Medicine. 2021;384(22):2124-2130. 36. Torjesen I. Covid-19: Risk of cerebral blood clots from disease is 10 times that from vaccination, study finds. BMJ. 2021;373:n1005. 37. Mezey É, Szalayova I, Hogden CT, et al. An immunohistochemical study of lymphatic elements in the human brain. Proceedings of the National Academy of Sciences. 2021;118(3):e2002574118. 38. Badolia R, Inamdar V, Manne BK, Dangelmaier C, Eble JA, Kunapuli SP. G(q) pathway regulates proximal C-type lectin-like receptor-2 (CLEC-2) signaling in platelets. Journal of Biological Chemistry. 2017;292(35):14516-14531. 39. Sung P-S, Huang T-F, Hsieh S-L. Extracellular vesicles from CLEC2-activated platelets enhance dengue virus-induced lethality via CLEC5A/TLR2. Nature Communications. 2019;10(1). 40. Ng H, Havervall S, Rosell A, et al. Circulating Markers of Neutrophil Extracellular Traps Are of Prognostic Value in Patients With COVID-19. Arteriosclerosis, Thrombosis, and Vascular Biology. 2021;41(2):988-994. 41. Welsh JD, Kahn ML, Sweet DT. Lymphovenous hemostasis and the role of platelets in regulating lymphatic flow and lymphatic vessel maturation. Blood. 2016;128(9):1169-1173. 42. Carey MA, Bradbury JA, Seubert JM, Langenbach R, Zeldin DC, Germolec DR. Contrasting Effects of Cyclooxygenase-1 (COX-1) and COX-2 Deficiency on the Host Response to Influenza A Viral Infection. The Journal of Immunology. 2005;175(10):6878-6884. 43. Kuhl PG, Bolds JM, Loyd JE, Snapper JR, FitzGerald GA. Thromboxane receptor-mediated bronchial and hemodynamic responses in ovine endotoxemia. American Journal of Physiology. 1988;254(2 Pt 2):R310-319. 44. Altavilla D, Canale P, Squadrito F, et al. Protective effects of BAY U 3405, a thromboxane A2 receptor antagonist, in endotoxin shock. Pharmacological Research. 1994;30(2):137-151. 45. Peters AT, Mutharasan RK. Aspirin for Prevention of Cardiovascular Disease. JAMA. 2020;323(7):676. 46. Chow JH, Khanna AK, Kethireddy S, et al. Aspirin Use Is Associated With Decreased Mechanical Ventilation, Intensive Care Unit Admission, and In-Hospital Mortality in Hospitalized Patients With Coronavirus Disease 2019. Anesthesia & Analgesia. 2021;132(4). 47. Saleh E, Moody MA, Walter EB. Effect of antipyretic analgesics on immune responses to vaccination. Human Vaccines & Immunotherapeutics. 2016;12(9):2391-2402.
Copyright: © 2021 The author(s). This is an open access article distributed under the terms of the Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited, a link to the Creative Commons license is provided, and any changes are indicated. The Creative Commons Public Domain Dedication waiver (https://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated. |