|
|
|
Free Neuropathology 2:5 (2021) |
|
Review |
|
Neurooncology: 2021 update |
|
Pieter Wesseling1,2 |
|
1 Department of Pathology, Amsterdam University Medical Centers, location VUmc, Brain Tumor Center Am-sterdam, Amsterdam, The Netherlands |
|
Corresponding author: |
|
Submitted: 26 February 2021 Accepted: 09 March 2021 Copyedited by: Deanna Fang Published: 17 March 2021 |
|
Keywords: Brain tumor, Molecular diagnostics, Glioma, Medulloblastoma, Tumor microenvironment, COVID-19 |
|
Abstract This article briefly presents 10 topics that were selected by the author as ‘top 10 discoveries’ published in 2020 in the broader field of neurooncological pathology including neurosciences as well as clinical neurooncology of interest for neurooncological pathology. The selected topics concern new information on the molecular characteristics of gliomas (infratentorial IDH-mutant diffuse astrocytomas, pediatric low-grade gliomas, infant-type high-grade gliomas, hypermutation in gliomas), the immunological aspects of the brain tumor microenvironment (TME), the impact of the TME on preclinical glioma models, and the importance of lymphatic drainage on brain tumor surveillance. Furthermore, important papers were published on two ‘new’ genetic syndromes predisposing to medulloblastoma, on liquid biopsy-based diagnosis of central nervous system (CNS) tumors, and on the ‘microbiome’ in glioblastomas (and other cancers). In the last part of this review, a dozen of papers are given as examples of papers that did not make it to the top 10 list of the author, underscoring the subjective component in the selection process. Acknowledging that 2020 will be remembered as the year in which the world changed because of the COVID-19 pandemic, some of the consequences of this pandemic for neurooncological pathology are briefly discussed as well. Hopefully, this review forms an incentive to appreciate the wealth of information provided by the papers that were used as building blocks for the present manuscript. Introduction Even though, in 2020, the COVID-19 pandemic had a quite dramatic, worldwide impact on the way we interacted (e.g. wearing of face masks, social distancing, travel restrictions, or even complete lockdowns), the publication of articles in international, peer-reviewed scientific journals seemed to continue more or less as usual. The author of this review was asked to contribute a review on the ‘top 10 discoveries’ in neurooncology published in 2020. As a search for candidate papers in PubMed using ‘brain & tumor & 2020’ or ‘spinal cord & tumor & 2020’ as key words resulted in over 10,000 hits, it took some time before the selection process for the ‘top ten discoveries’ was finished. Here follows the list of the discoveries that emerged because they can be expected to have substantial implications for neurooncological pathology:
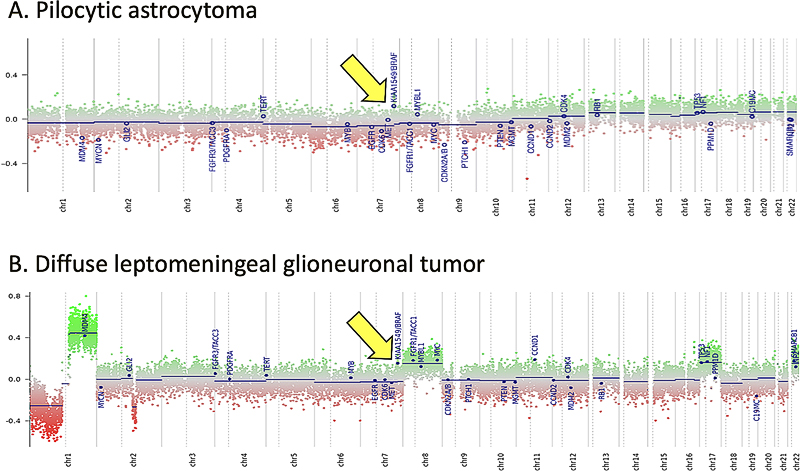
The order in this list is not based on any ranked importance of the findings but reflects an attempt to create some flow in this review. Furthermore, acknowledging that authors did their best to optimally summarize the essence of their findings in (especially the abstracts of) the papers, in this review several phrases were copied and pasted from the original papers, always accompanied by adding the reference. Of course, there is quite a subjective component in the selection of papers presented in this review. For example, the selection was not per se solely based on articles in the high(est)-impact journals but also aimed for a somewhat broader blend of topics. Ergo, other colleagues would probably have selected other papers, but, as far as I am concerned, that is all in the game (‘de gustibus non est disputandum’). To make up for this situation to some degree, a few papers that just did not make it to my top 10 list will be briefly mentioned in the Discussion. Because of the extraordinary impact of the COVID-19 pandemic in 2020 on our (professional) lives, some COVID-19 aspects related to neurooncological pathology are very briefly discussed as well. Topic 1. Infratentorial IDH-mutant astrocytomas are different [1] In the World Health Organization (WHO) 2016 classification, three main groups of adult-type diffuse gliomas are listed: astrocytomas, IDH-mutant; oligodendrogliomas, IDH-mutant and 1p/19q-codeleted; and astrocytomas, IDH-wildtype (with glioblastoma IDH-wildtype as by far the most frequent representative in this last group) [15]. In the upcoming 5th edition of the WHO classification (to be published in 2021), some changes can be expected in the definitions and nomenclature of IDH-mutant astrocytomas. This is in line with what has been proposed by different updates of the consortium to Improve Molecular and Practical Approaches to CNS tumor Taxonomy (cIMPACT-NOW). cIMPACT-NOW update 5 suggested to grade the IDH-mutant astrocytomas as WHO grade IV if molecular analysis reveals the presence of homozygous CDKN2A/B deletion and not only if the tumor histologically shows features of the highest malignancy grade [16]. Furthermore, cIMPACT-NOW update 6 proposed to group the WHO grade II-IV diffuse IDH-mutant astrocytic tumors under a single name (“Astrocytoma, IDH mutant”) with Arabic numerals assigned for the grade. The most malignant form in this group would then be ‘Astrocytoma, IDH-mutant, grade 4’ (rather than ‘Glioblastoma, IDH-mutant’), facilitating discrimination from the much more frequent and more aggressive ‘IDH–wildtype glioblastoma’ [17]. In these considerations, the location of the IDH-mutant astrocytic tumors is not taken into account. Indeed, in the vast majority of patients presenting with a diffuse IDH-mutant astrocytoma, the tumor is located in a cerebral hemisphere. Banan R et al (with Hartmann C and Reuss D as corresponding authors) published in Acta Neuropathologica the clinical and pathological characteristics of a series of 42 primary infratentorial IDH-mutant astrocytic tumors [1]. The mean age of the patients in this series was 37 years (similar to the patients with supratentorial tumors), the male:female ratio was 1.8:1, and approximately 30%, 40% and 30% were considered to be WHO grade II, III and IV, respectively. Interestingly, only 1 out of 4 of these infratentorial IDH-mutant tumors had the IDH1 R132H mutation (in supratentorial tumors this is > 80%), with IDH1 R132C/G and IDH2 R172S/G as the most frequent ‘other’ IDH mutations. Also, ATRX loss and MGMT promoter methylation were significantly less frequently found in the infratentorial compartment. Of note, gene panel sequencing revealed two samples with IDH1 R132C/H3F3A K27M co-mutations in this series. Overall, the clinical outcomes of patients with infratentorial IDH-mutant astrocytomas were significantly better than for patients with diffuse midline gliomas, H3K27M-mutant but were significantly worse than for patients with supratentorial IDH-mutant astrocytomas when the two cases with IDH1/H3K27M co-mutation were included. After exclusion of these two cases, only a trend toward worse outcome for infratentorial IDH-mutant astrocytomas was seen. This study shows that diffuse gliomas in the brainstem and cerebellum are easily missed as being IDH-mutant in cases where only immunohistochemistry for IDH1 R132H mutant protein is performed and that H3K27M mutations co-occur in a small subset of these tumors and signify worse prognosis. Topic 2. Molecular landscape of pediatric low-grade gliomas [2] Ryall S et al (with Tabori U and Hawkins C as co-senior authors) published a paper in Cancer Cell reporting the results of combined clinical, morphological and molecular profiling of > 1000 well-annotated, pediatric low-grade gliomas (pLGGs) with extensive clinical follow-up. The vast majority of the cases that could be adequately analyzed at the molecular level were found to harbor a driver alteration leading to activation of the RAS/MAPK pathway, while those without an identified alteration often showed upregulation of this pathway as well. Based on the type of molecular alteration, pLGGs could be broadly classified in different classes: rearrangement-driven/fusion-positive tumors versus tumors with a single-nucleotide variant (SNV). Tumors in the former category were diagnosed at a younger age, enriched for WHO grade I histology, progressed infrequently, and rarely resulted in death as compared to tumors in the latter category. Correlation of type of molecular alteration with outcome (incidence of disease progression or death) allowed for stratification of pLGGs into three risk categories: low risk (BRAF fusions, NF1 alterations, FGFR2 fusions, MYB/MYBL1 rearrangements, FGFR1 tyrosine kinase duplications, FGFR1-TACC1 fusion); intermediate risk (BRAF p.V600E, IDH1 p.R132H, FGFR1 SNV, MET SNV); and high risk (BRAF p.V600E + CDKN2A deletion, H3.3 p.K27M). Additionally, a small, remaining group of ‘unknown risk’ was found with fusions of or SNVs in other genes. Importantly, while most pLGGs in children with the genetic pre-disposition disorder NF1 occurred as optic pathway glioma and had a good prognosis, NF1 pLGGs arising outside the optic pathway had significantly worse overall and progression-free survival. Also, the most common KIAA1549-BRAF fusion, i.e. 16:09 (involving exon 16 of KIAA1549 and exon 9 of BRAF), was found to be associated with cerebellar location and good prognosis. In contrast, the KIAA1549-BRAF 15:09 fusion was the only fusion type seen in hemispheric tumors, the primary fusion found in tumors with ‘extensive dissemination’, and associated with a worse progression-free survival. These findings indeed provide very helpful guidance for the prognostication and clinical management of patients with pLGGs (e.g. ‘watch & wait’ for low-risk tumors versus a more aggressive approach for intermediate and high-risk tumors). Obviously, for individual patients, such information should be evaluated in the context of the clinical situation, other molecular findings, and the exact WHO diagnosis (Figure 1).
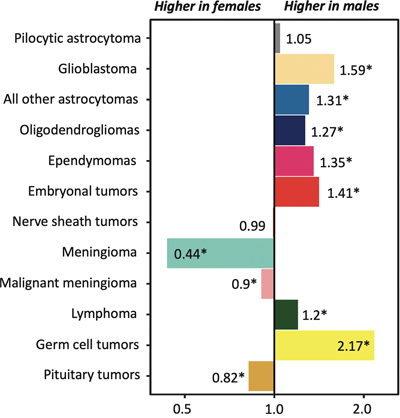
Fig. 1. KIAA-BRAF1549 duplication (and fusion) as seen in the copy number variation (CNV)-profile of a pilocytic astrocytoma (A) and a diffuse leptomeningeal glioneuronal tumor (DLGNT) (B). A tandem duplication of chromosome 7q34, indicative of the KIAA1549-BRAF fusion, is a frequent event in pilocytic astrocytomas and DLGNTs. Such a duplication is generally clearly visible as a relatively high position for this region (indicated by arrows in A and B) compared to the rest of chromosome 7 in the detailed CNV profiles that can be obtained using e.g. methylome profiling [55]. Ryall S et al propose disease stratification of pediatric low-grade gliomas (pLGGs) based on the type of molecular alteration(s) in the tumor, with most KIAA1549-BRAF fusion-positive tumors belonging to the low-risk category [2]. It is important to put such information in perspective of clinical and other molecular findings and of the WHO diagnosis of the individual patient. For example, all DLGNTs typically show loss of chromosome 1p. The CNV profile of the DLGNT depicted in B additionally shows gain of chromosome arm 1q and of chromosome 8, two features that fit very well with the more aggressive ‘methylation class 2’ of these tumors [56]. Interestingly, RT-PCR analysis of this DLGNT revealed the 15:09 KIAA1549-BRAF fusion, i.e. the fusion that, according to Ryall S et al, is the primary one seen in pLGGs with ‘extensive dissemination’ and that is associated with worse progression-free survival. Topic 3. Gene fusions in infant-type high-grade gliomas [3] In 2019, Guerreiro Stucklin AS, Ryall S, et al (with Tabori U and Hawkins C as last authors) published a study of 171 glioma samples from 150 infants in Nature Communications in which they identified three main subgroups of infant gliomas: 1) hemispheric RTK-driven tumors, including those showing fusion involving ALK, ROS1, NTRK, or MET, enriched for high-grade gliomas and with an intermediate clinical outcome; 2) hemispheric RAS/MAPK-driven tumors, characterized by excellent long-term survival post-surgery; and 3) midline RAS/MAPK-driven tumors, enriched for low-grade glioma with BRAF alterations and a relatively poor outcome, even after conventional chemotherapeutic approaches [18]. In 2020, Clarke M, Mackay A, Ismer B, et al (last author Jones C) published in Cancer Discovery the findings of a thorough histological and molecular analysis of 241 tumors diagnosed as high-grade and/or diffuse gliomas in children < 4 years of age at the time of diagnosis [3]. After excluding non-gliomas and gliomas matching known subtypes, 130 infant gliomas remained that seemed to be part of an "intrinsic" spectrum of disease specific to the infant population. These included those with targetable MAPK pathway alterations. A large proportion of remaining cases harbored gene fusions; these tumors typically occurred in very young children (age at diagnosis less than < 1 year), were located in the cerebral hemispheres, and (like the ‘group 1 tumors’ as reported by Guerreiro Stucklin AS et al) frequently carried fusions involving ALK (n = 31), NTRK1/NTRK2/NTRK3 (n = 21), ROS1 (n = 9), or MET (n = 4) as their driving alterations. Importantly, compared to the fusion-negative tumors in this group, these fusion-positive tumors had significantly better outcomes. Also, they would be good candidates for targeted therapy. This study is a next example of how in-depth molecular profiling of pediatric brain tumors not only greatly helps to improve prognostication for the children suffering from these neoplasms, but also to discover actionable targets that have the potential to substantially improve their prognosis [19]. Topic 4. Hypermutation in gliomas [4] While only a limited number of newly diagnosed gliomas are characterized by an inherited mismatch repair (MMR) defect and/or a ‘hypermutator’ phenotype, recurrent gliomas more often show such a phenotype, especially after alkylating chemotherapy [20]. The combination of low tumor mutational burden (TMB) and a highly immunosuppressive microenvironment in most newly diagnosed diffuse gliomas renders (at least so far) effective immunotherapy for these tumors quite challenging. Assuming that the hypermutator status leads to an increased expression of neoantigens, gliomas/glioblastomas with a hypermutator phenotype could be better candidates for immune checkpoint blockade. In a paper in Nature, Touat M and Li YY, et al (with Beroukhim R, Bandopadhayay P, Bielle F, and Ligon KL as supervising authors) report the results of their comprehensive analysis of the molecular determinants of mutational burden and signatures in 10,294 gliomas [4]. The gliomas were classified into molecular subgroups according to histopathology as well as IDH1/IDH2 and 1p/19q codeletion status. The median TMB in all samples was 2.6 mutations per Mb (range 0.0–781.3). 558 (5.4%) tumors that were designated as being hypermutated (median TMB 50.8 mutations per Mb, range 8.8–781.3) were analyzed more in depth. The majority of de novo hypermutated gliomas harbored mutational signatures associated with defects in the MMR pathway (COSMIC signatures 6, 15, 26 and 14) or the DNA polymerase POLE (10 and 14) (2/3 and 1/3 of the samples, respectively), implying that constitutional deficiency in MMR or POLE was likely to be the underlying genetic cause of hypermutation. In contrast, almost all gliomas showing high TMB post-treatment had a mutational signature associated with temozolomide exposure (signature 11), and half of these samples showed a co-existing minor MMR- or POLE-deficiency signature component, suggesting that defective DNA repair and mutagen exposure cooperate to drive hypermutation in these recurrent gliomas. The observation that bulk analyses of such post-treatment hypermutated glioma did not readily detect microsatellite instabilities (MSIs), while single-cell whole-genome sequencing analysis did identify microsatellite mutations, can be explained by intra-tumor heterogeneity for this characteristic and a lack of sufficient evolutionary time to select subclonal MSI populations. While the therapy-induced SNV mutations might not readily elicit effective antitumor responses (because of, for example, the quality of the mutations and/or the subclonal nature of their associated neoantigens), longer treatment exposure or combinatorial strategies may improve the efficacy of checkpoint blockade for such tumors. Importantly, and acknowledging that acquired MMR deficiency occurs in the tumors that are considered as being the most temozolomide-sensitive, it is not yet clear whether the acquired MMR deficiency outweighs the positive effects of temozolomide in gliomas. The finding that MMR-deficient cells retain sensitivity to CCNU supports the hypothesis that hypermutation reduces cellular fitness and tolerance to DNA-damaging agents other than temozolomide. One of the conclusions of the authors is that longitudinal, molecular analysis of diffuse gliomas can help improve therapeutic management and, ultimately, prognosis as well. Topic 5. Immunological aspects of the brain tumor microenvironment [5-7] In two papers published back-to-back in Cell in June 2020, the authors report on the abundance and heterogeneity of tissue-resident and peripherally recruited leucocytes in glial and metastatic brain tumors [5, 6]. Klemm F et al (last author Joyce J [5]) comprehensively characterized the tumor microenvironment (TME) of gliomas and brain metastases using flow cytometry, RNA sequencing, protein arrays, culture assays, and spatial tissue characterization. They found that ‘education’ of immune cell types in the TME depends on tumor origin and IDH mutational status, with pronounced differences in proportional abundance of tissue-resident microglia, infiltrating monocyte-derived macrophages, neutrophils, and T cells. Friebel B, Kapolou K, et al (last authors Neidert MC, Becher B [6]) mapped the leukocyte landscape of brain tumors using high-dimensional single-cell profiling (CyTOF) and found a heterogeneous composition of tissue-resident and invading immune cells within the TME, allowing for a clear distinction between gliomas and brain metastases. Gliomas typically showed tissue-resident, reactive microglia, whereas tissue-invading leukocytes accumulated in brain metastases. Tissue-invading tumor-associated macrophages showed a distinctive signature trajectory, revealing tumor-driven instruction along with contrasting lymphocyte activation and exhaustion. Indeed, these integrated analyses further elucidate the multifaceted immune cell activation within brain tumors and are instrumental for a rational design of more efficacious, targeted immunotherapy strategies. In the study published in Cancer Discovery, Bayik D et al (last author Lathia JD) used mouse models to study the presence of monocytic versus granulocytic myeloid-derived suppressor cells (mMDSCs/gMDSCs) [7]. MDSCs are known to be elevated in blood and tumor tissue of patients with glioblastoma and for blocking antitumor immunity. Bayik D et al observed that, in their models, mMDSCs were enriched in the male tumors, whereas gMDSCs were elevated in the blood of females. Furthermore, depletion of gMDSCs extended survival only in female mice. Using gene-expression signatures coupled with network medicine analysis, the authors demonstrated that mMDSCs could be targeted with antiproliferative agents in males, whereas gMDSC function could be inhibited by IL1β blockade in females. Analysis of patient data confirmed that proliferating mMDSCs were predominant in male tumors and that a high gMDSC/IL1β gene signature correlated with poor prognosis in female patients. Interestingly, there is a quite striking sex disparity for glioblastomas at other levels as well. For example, the incidence of glioblastoma is significantly higher in males than in females (incidence rate ratio 1.59:1) (Figure 2), and there is a survival advantage for females with glioblastoma independent of treatment, age, Karnofsky Performance Status (KPS), or IDH mutation status [21, 22]. The work of Bayik et al may explain some of such sex differences in patients with glioblastoma and indicate that (immuno)therapeutic approaches should be adapted to the patient’s gender.
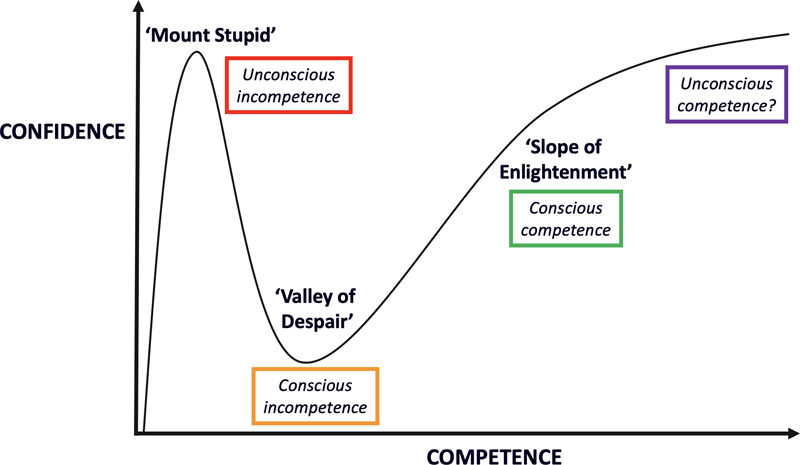
Fig. 2. Incidence rate ratios by sex for primary brain tumors. The incidence of most gliomas and embryonal brain tumors is significantly higher in males. For glioblastomas the male:female ratio is 1.59:1 [21]. Furthermore, there is a female survival advantage for patients with glioblastoma independent of treatment, age, Karnofsky Performance Status (KPS), or IDH mutation status [22]. The work of Bayik et al may provide support for the hypothesis that differences in immune system functions contribute to these sex differences and that (immuno)therapeutic approaches should be adapted to the patient’s gender. Figure based on Ostrom QT et al (2020) [21]. * = p<0.05. Topic 6. Lymphatic drainage enables brain tumor surveillance [8, 9] The central nervous system (CNS) has long been considered as lacking lymphatic drainage, a situation which could then contribute to the generally limited immunological response in glioblastomas and other brain tumors. Later on, however, it became clear that lymphatic vessels are present in the intracranial meninges along the dural sinuses. These lymphatics drain brain-derived soluble waste to deep cervical lymph nodes, thereby directly connecting the brain with the peripheral immune system. Recent studies also demonstrated the presence of meningeal lymphatics along the spinal cord and even suggested that the cerebrospinal fluid (CSF) is primarily drained via lymphatics, rather than into the dural sinuses [23]. In a study published in Nature, Song E et al (last authors Thomas JL and Iwasaki A) report that in mice orthotopically injected with glioblastoma cells, prophylactic injection of the lymphangiogenesis-promoting protein VEGFC in the CSF resulted in enhanced priming of CD8 T cells in the draining deep cervical lymph nodes, migration of such cells into the tumor, rapid clearance of the glioblastoma and a long-lasting anti-tumor memory response [8]. Transfection of a VEGFC-expression mRNA construct was found to work synergistically with checkpoint blockade therapy to eradicate existing glioblastoma. Furthermore, Hu X, Deng Q, Ma L, et al (last author Luo J) also published a study last year on the role of meningeal lymphatic vessels in mice with intracranial gliomas or metastatic melanomas [9]. Disruption of dorsal meningeal lymphatics alone impaired the dissemination of tumor cells and dendritic cells from the brain tumors to deep cervical lymph nodes, while such trafficking of dendritic cells was found to be increased in mice with enhanced dorsal meningeal lymphangiogenesis. Also, disruption of dorsal meningeal lymphatics alone (without affecting basal or nasal meningeal lymphatics) significantly reduced the efficacy of combined anti-PD-1/CTLA-4 checkpoint therapy in striatal tumor models, while tumors overexpressing VEGFC displayed a better response to such combination therapy. These studies suggest that the immunosurveillance-promoting capacity of VEGFC may be exploited to increase the efficacy of immunotherapeutic approaches for brain tumors. Topic 7. Impact of tumor microenvironment on preclinical glioblastoma models [10] A wide variety of preclinical models for human glioblastomas serve as an important tool for studying these brain tumors. However, no model is perfect, and it is important to determine if/how different models recapitulate different aspects of human glioblastomas. To address this issue, Pine AR, Cirigliano SM, et al (last author Fine HA) performed a complete transcriptomic characterization of tumor cells from 5 patients across four patient-specific glioblastoma–derived model types: 1) glioma spheres (GSs), 2) tumor organoids (TOs), 3) glioblastoma cerebral organoids (GLICOs), and 4) patient-derived xenografts (PDXs). They found that GLICOs and PDXs more closely recapitulated the invasive growth of human glioblastomas. Also, the results obtained with bulk and single cell RNA sequencing analysis of these models more closely resembled the human tumors. Compared to the other models, GLICOs were enriched for a neural progenitor-like cell subpopulation, showed lower percentages of mesenchymal cells, showed retention of neural and oligodendrocyte progenitor cell populations and recapitulated the cellular states and their plasticity found in the corresponding human tumors best. Although GLICOs thus had a profile most similar to that of the parental glioblastomas, the similarity between GLICOs and the original glioblastomas was reduced when GLICO cells were replated in two-dimensional culture conditions. This work underscores the critical impact of the microenvironment in glioblastoma models on the degree to which cellular states, as found in human glioblastomas, are recapitulated. Thereby, this study serves as a kind of reality check for those who exploit such preclinical models for elucidation of the pathobiology of and identification of therapeutic approaches for glioblastomas in clinical practice. Topic 8. Newly discovered medulloblastoma predisposition syndromes [11, 12] Medulloblastomas are by far the most frequent of the embryonal CNS tumors, most commonly present in childhood and display considerable biological heterogeneity, with distinct molecularly defined groups listed in the WHO classification. A recent review reports that pathogenic germline variants in established cancer predisposition genes can be identified in about 5% of patients with medulloblastoma [24]. This includes syndromes with a germline defect in SUFU or PTCH1 (nevoid basal cell carcinoma syndrome/Gorlin syndrome), TP53 (Li–Fraumeni syndrome), APC (familial adenomatous polyposis), CREBBP (Rubinstein–Taybi syndrome), NBS1 (Nijmegen breakage syndrome), PALB2, and BRCA2. In 2020, two additional germline mutations were reported that predispose to pediatric medulloblastoma. Waszak SM, Robinson GW, et al (last authors Korbel JO, Northcott PA and Pfister SM) published a study in Nature of 1022 patients with medulloblastoma from whom blood samples and tumor samples were analyzed for germline mutations in 110 cancer predisposition genes [11]. They identified rare germline loss-of-function variants across the Elongator Complex Protein 1 (ELP1) gene in 14% of pediatric patients with the Sonic Hedgehog medulloblastoma subgroup (MBSHH). ELP1 was the most common medulloblastoma predisposition gene and increased the prevalence of genetic predisposition to 40% among pediatric patients in this MBSHH subgroup. Parent-offspring and pedigree analyses identified two families with a history of pediatric medulloblastoma. Most ELP1-associated medulloblastomas also exhibited somatic alterations in PTCH1. Tumors from patients with ELP1-associated MBSHH were characterized by universal biallelic inactivation of ELP1 owing to somatic loss of chromosome arm 9q alterations. The tumor cells were found to show changes consistent with loss of protein homeostasis due to Elongator deficiency. The authors conclude that such a genetic predisposition to proteome instability may be a determinant in the pathogenesis of other (pediatric) cancers as well and may provide new targets for therapeutic interference. In another study, Begemann M, Waszak SM, et al (last authors Pfister SM, Kontny U, Kurth I, and published in the Journal of Clinical Oncology) investigated families with childhood medulloblastoma to identify predisposing germline mutations [12]. Initial findings were extended to genomes and epigenomes of 1,044 medulloblastoma cases. The authors identified in six patients with infant-onset medulloblastoma a heterozygous germline mutation in the G protein-coupled receptor 161 (GPR161) gene, which is located on chromosome 1q. GPR161 mutations were exclusively associated with the MBSHH subgroup and accounted for 5% of infant cases in these cohorts. Molecular profiling revealed loss of heterozygosity (LOH) at GPR161 in all affected MBSHH tumors without additional somatic driver events. Additionally, analysis of 226 MBSHH tumors revealed somatic copy-neutral LOH of chromosome 1q as a characteristic hallmark and the primary mechanism for biallelic inactivation of GPR161 in the affected MBSHH tumors. Obviously, the diagnosis of an underlying predisposition syndrome as reported in these papers is important because of its implications for the management of the patients and their families. Topic 9. Liquid biopsy diagnosis of CNS tumors [13] Circulating tumor DNA (ctDNA) in blood is considered to be an easily accessible source of diagnostic, prognostic and/or predictive information that may be very helpful for improving the management of cancer patients. So far, however, for patients with primary brain tumors, the use of CSF appeared to be a more promising biosource because of the limited abundance of ctDNA in blood of these patients. At the same time, approaches that go beyond DNA sequence information, such as analysis of epigenetic signatures in ctDNA, can help to boost the use of blood for liquid biopsy diagnostics [25]. In the study published in Nature Medicine, Nassiri F, Chakravarthy A, Feng S, et al (last authors Zadeh G and De Carvalho DD) used cell-free methylated DNA immunoprecipitation and high-throughput sequencing (cfMeDIP-seq) to recover and profile methylated DNA fragments from plasma [13]. Plasma samples of patients with diffuse (IDH-wildtype and IDH-mutant) gliomas, of patients with other intracranial and extracranial tumors, and of healthy controls were used. These samples were split into training and test sets, and random forest classifiers were trained using the top 300 differentially methylated regions for the different tumor classes. Indeed, the classifier was able to accurately distinguish different tumors based on the differences in DNA methylation profiles. The potential of such a methylome-based liquid biopsy approach has also already been reported for non-CNS tumors and can be applied to other biosources (e.g. urine) as well [26, 27], underscoring the potential of analysis of methylated plasma ctDNA to facilitate blood-based diagnosis and monitoring of patients with cancer. Topic 10. The glioblastoma microbiome [14] Tumor formation involves the co-evolution of neoplastic cells together with extracellular matrix, tumor vasculature and immune cells. Evaluating tumors as complete ‘organs’, and not simply as masses of transformed tumor cells, is of paramount importance because heterologous cell types within tumors can actively influence therapeutic response [28]. Bacteria were first detected in human tumors more than 100 years ago, raising the possibility that the tumor microbiome may be an additional player in the complex tumor ecosystem. However, the existence of the tumor microbiome remained an issue of debate because of the suspicion of sample contamination. In their study published in Science, Nejman D, Livyatan I, Fuks G, et al (last authors Shental N, Straussman R) characterized the microbiome of 1010 tumor samples from seven human cancer types (breast, lung, ovary, pancreas, melanoma, bone, and brain/glioblastoma) as well as of 516 normal samples (including normal tissue adjacent to the tumor from the same patients) [14]. The authors took multiple measures to minimize and control for contamination. They used PCR sequencing techniques to gain species-level resolution as well as multiple visualization methods and ‘culturomics’ for identification of bacteria. The authors report the presence of bacteria in each tumor type, including in tumors that have no direct connection with the external environment such as glioblastomas. The detection rate ranged from 14.3% in melanomas to >60% in breast, pancreatic, and bone tumors. Breast tumors had a richer and more diverse microbiome than all other tumor types tested, and live bacteria could be cultured from these tumors. The bacteria were found to be predominantly localized intracellularly in both cancer cells and immune cells, with different tumor types showing a distinct microbiome composition. The authors also noted correlations between intratumor bacteria and the smoking status and/or the response to immunotherapy of the patients. Obviously, these findings immediately raise additional questions, for example: What (if any) is the role of intratumoral bacteria in the development of cancer? And how does the tumor microbiome affect the immune tumor microenvironment and the response to immune therapy? The answers to such questions are crucial for further elucidation of the existence and the importance of the tumor microbiome in cancer [29]. Discussion Hopefully, this review indeed provides concise, easily digestible information on the topics selected by the author as the top 10 discoveries in the year 2020. Of course, during the selection process, quite a few papers were noted that didn’t make it to the top 10 list but that are certainly of great interest as well. For example, it is reassuring to learn that the use of molecular markers for the diagnosis of glioblastoma, IDH-wildtype as suggested by cIMPACT-NOW update 3 was corroborated in an independent study [30, 31], but it is also good to know that some caution may be warranted for histologically low-grade IDH-wildtype diffuse astrocytic tumors with an isolated TERT promoter mutation [32]. Furthermore, some new tumor types were recognized that will make it to the new WHO classification of CNS tumors (especially diffuse glioneuronal tumor with oligodendroglioma-like features and nuclear clusters (DGONC) and desmoplastic myxoid tumor of the pineal region, SMARCB1-mutant [33, 34]), and new insights were obtained in areas such as the oncogenesis of H3.3G34-mutant gliomas [35], posterior fossa group A (PFA) ependymomas [36], subependymal giant cell astrocytomas [37], and in the methylation characteristics of germline-driven replication repair-deficient high-grade gliomas (‘unique hypomethylation patterns’) [38]. Very interesting new information was published on how glioblastomas interact with neurons [39], and a roadmap was presented for shaping the emerging field of Cancer Neuroscience [40]. In depth (epi)genetic analysis of single (brain tumor) cells is ‘hot’ nowadays and indeed provides an enormous amount of new information. A novel tool for exploiting single-cell RNA sequencing data, the single-cell Tumor-Host Interaction tool (scTHI), was published in a journal with the impressive name Gigascience [41]. Although not included in the top 10 list in this review, much has been discovered in 2020 about and because of the COVID-19 pandemic, with some consequences for neurooncology as well. Because of the very infectious and often serious character of the disease, governments had to take drastic actions such as enforcing the wearing of face masks, installing social distancing measures and limiting the free movement of citizens. Combined with the finding that cancer patients have an increased risk of dying because of COVID-19 infection, this has caused increased stress and anxiety amongst brain tumor patients and their caregivers [42, 43]. Neurooncologic care needed to be delivered in an adapted form, minimizing exposures for patients and addressing risks and benefits of all therapeutic interventions, while at the same time ensuring the health of the multi-disciplinary neurooncology workforce as much as possible [44]. Interestingly, it appeared that some diagnostic (neuro)pathology services can be reliably provided while working from home when using platforms for whole slide images (WSIs). Of course, some activities (e.g. tissue sampling, processing, cutting, staining, molecular analyses) still need to be done in the institutes [45-47]. As an emergency response to the pandemic, the UK Royal College of Pathologists provided guidance for remote reporting of digital pathology slides and the US Food and Drug Administration (FDA) granted a waiver for the use of readily available consumer monitors at home [48, 49]. Furthermore, the COVID-19 pandemic has boosted the exploitation of present-day technology for teaching of medical students and pathology residents (e.g. using digital lectures, videoconferencing, and online resources utilizing collections of WSIs) [50, 51] and has drastically changed the way scientific communities interact now that meetings cannot be organized in their traditional format. However, the ‘human touch’ is easily missed in such virtual encounters. Finding the right balance between attending web-based meetings, getting (the rest of) our daily job done from home, and maintaining a healthy personal life can be challenging [52]. Also, it remains to be seen what the long-term impact of the pandemic is on cancer research [53]. Last but not least, ignorance of one’s own ignorance is worrisome, especially so because not infrequently it is combined with overconfidence, a phenomenon called the Dunning-Kruger effect [54]. Improving skills and metacognitive competence helps one to move on from conscious incompetence to a phase of conscious competence that is more in balance with the level of confidence (Figure 3). Although a review like this only allows for ‘scratching the surface’ of the selected discoveries, it hopefully does help one to move further away from unconscious incompetence, especially so if the review indeed works as an incentive to read and appreciate the original papers that were used as building blocks for the present manuscript.
Fig. 3. The Dunning-Kruger effect: A remarkable relationship between competence and confidence. In 1999, David Dunning and Justin Kruger described the phenomenon that people tend to initially hold overly favorable views of their abilities in many social and intellectual domains [54]. Indeed, people not infrequently share strong opinions on particular topics without being hindered by their lack of competence (in other words, they’re on top of ‘Mount Stupid’ without knowing it). Improving skills and metacognitive competence helps to move on via the ‘Valley of Despair’ (conscious incompetence: knowing that you only have very limited knowledge and/or understanding of a particular topic) to the ‘Slope of Enlightenment’, where competence and confidence are more in balance. Acknowledging that competence is often a matter of continuing education and professional development, I doubt if the ultimate goal in medicine is really to strive for the level of unconscious competence (hence the question mark behind that term). Meanwhile, this review hopefully helps to climb the ‘Slope of Enlightenment’ somewhat further. References 1. Banan, R., et al., Infratentorial IDH-mutant astrocytoma is a distinct subtype. Acta Neuropathol, 2020. 140(4): p. 569-581. 2. Ryall, S., et al., Integrated Molecular and Clinical Analysis of 1,000 Pediatric Low-Grade Gliomas. Cancer Cell, 2020. 37(4): p. 569-583 e5. 3. Clarke, M., et al., Infant High-Grade Gliomas Comprise Multiple Subgroups Characterized by Novel Targetable Gene Fusions and Favorable Outcomes. Cancer Discov, 2020. 10(7): p. 942-963. 4. Touat, M., et al., Mechanisms and therapeutic implications of hypermutation in gliomas. Nature, 2020. 580(7804): p. 517-523. 5. Klemm, F., et al., Interrogation of the Microenvironmental Landscape in Brain Tumors Reveals Disease-Specific Alterations of Immune Cells. Cell, 2020. 181(7): p. 1643-1660 e17. 6. Friebel, E., et al., Single-Cell Mapping of Human Brain Cancer Reveals Tumor-Specific Instruction of Tissue-Invading Leukocytes. Cell, 2020. 181(7): p. 1626-1642 e20. 7. Bayik, D., et al., Myeloid-Derived Suppressor Cell Subsets Drive Glioblastoma Growth in a Sex-Specific Manner. Cancer Discov, 2020. 10(8): p. 1210-1225. 8. Song, E., et al., VEGF-C-driven lymphatic drainage enables immunosurveillance of brain tumours. Nature, 2020. 577(7792): p. 689-694. 9. Hu, X., et al., Meningeal lymphatic vessels regulate brain tumor drainage and immunity. Cell Res, 2020. 30(3): p. 229-243. 10. Pine, A.R., et al., Tumor Microenvironment Is Critical for the Maintenance of Cellular States Found in Primary Glioblastomas. Cancer Discov, 2020. 10(7): p. 964-979. 11. Waszak, S.M., et al., Germline Elongator mutations in Sonic Hedgehog medulloblastoma. Nature, 2020. 580(7803): p. 396-401. 12. Begemann, M., et al., Germline GPR161 Mutations Predispose to Pediatric Medulloblastoma. J Clin Oncol, 2020. 38(1): p. 43-50. 13. Nassiri, F., et al., Detection and discrimination of intracranial tumors using plasma cell-free DNA methylomes. Nat Med, 2020. 26(7): p. 1044-1047. 14. Nejman, D., et al., The human tumor microbiome is composed of tumor type-specific intracellular bacteria. Science, 2020. 368(6494): p. 973-980. 15. Wesseling, P. and D. Capper, WHO 2016 Classification of gliomas. Neuropathol Appl Neurobiol, 2018. 44(2): p. 139-150. 16. Brat, D.J., et al., cIMPACT-NOW update 5: recommended grading criteria and terminologies for IDH-mutant astrocytomas. Acta Neuropathol, 2020. 139(3): p. 603-608. 17. Louis, D.N., et al., cIMPACT-NOW update 6: new entity and diagnostic principle recommendations of the cIMPACT-Utrecht meeting on future CNS tumor classification and grading. Brain Pathol, 2020. 30(4): p. 844-856. 18. Guerreiro Stucklin, A.S., et al., Alterations in ALK/ROS1/NTRK/MET drive a group of infantile hemispheric gliomas. Nat Commun, 2019. 10(1): p. 4343. 19. Wong, M., et al., Whole genome, transcriptome and methylome profiling enhances actionable target discovery in high-risk pediatric cancer. Nat Med, 2020. 26(11): p. 1742-1753. 20. Barthel, F.P., et al., Longitudinal molecular trajectories of diffuse glioma in adults. Nature, 2019. 576(7785): p. 112-120. 21. Ostrom, Q.T., et al., CBTRUS Statistical Report: Primary Brain and Other Central Nervous System Tumors Diagnosed in the United States in 2013-2017. Neuro Oncol, 2020. 22(12 Suppl 2): p. iv1-iv96. 22. Ostrom, Q.T., et al., Females have the survival advantage in glioblastoma. Neuro Oncol, 2018. 20(4): p. 576-577. 23. Oliver, G., et al., The Lymphatic Vasculature in the 21(st) Century: Novel Functional Roles in Homeostasis and Disease. Cell, 2020. 182(2): p. 270-296. 24. Waszak, S.M., et al., Spectrum and prevalence of genetic predisposition in medulloblastoma: a retrospective genetic study and prospective validation in a clinical trial cohort. Lancet Oncol, 2018. 19(6): p. 785-798. 25. van der Pol, Y. and F. Mouliere, Toward the Early Detection of Cancer by Decoding the Epigenetic and Environmental Fingerprints of Cell-Free DNA. Cancer Cell, 2019. 36(4): p. 350-368. 26. Shen, S.Y., et al., Sensitive tumour detection and classification using plasma cell-free DNA methylomes. Nature, 2018. 563(7732): p. 579-583. 27. Nuzzo, P.V., et al., Detection of renal cell carcinoma using plasma and urine cell-free DNA methylomes. Nat Med, 2020. 26(7): p. 1041-1043. 28. Junttila, M.R. and F.J. de Sauvage, Influence of tumour micro-environment heterogeneity on therapeutic response. Nature, 2013. 501(7467): p. 346-54. 29. Atreya, C.E. and P.J. Turnbaugh, Probing the tumor micro(b)environment. Science, 2020. 368(6494): p. 938-939. 30. Brat, D.J., et al., cIMPACT-NOW update 3: recommended diagnostic criteria for "Diffuse astrocytic glioma, IDH-wildtype, with molecular features of glioblastoma, WHO grade IV". Acta Neuropathol, 2018. 136(5): p. 805-810. 31. Tesileanu, C.M.S., et al., Survival of diffuse astrocytic glioma, IDH1/2 wildtype, with molecular features of glioblastoma, WHO grade IV: a confirmation of the cIMPACT-NOW criteria. Neuro Oncol, 2020. 22(4): p. 515-523. 32. Berzero, G., et al., IDH-wildtype lower grade diffuse gliomas: the importance of histological grade and molecular assessment for prognostic stratification. Neuro Oncol, 2020. 33. Deng, M.Y., et al., Diffuse glioneuronal tumour with oligodendroglioma-like features and nuclear clusters (DGONC) - a molecularly defined glioneuronal CNS tumour class displaying recurrent monosomy 14. Neuropathol Appl Neurobiol, 2020. 46(5): p. 422-430. 34. Thomas, C., et al., Desmoplastic myxoid tumor, SMARCB1-mutant: clinical, histopathological and molecular characterization of a pineal region tumor encountered in adolescents and adults. Acta Neuropathol, 2020. 139(2): p. 277-286. 35. Chen, C.C.L., et al., Histone H3.3G34-Mutant Interneuron Progenitors Co-opt PDGFRA for Gliomagenesis. Cell, 2020. 183(6): p. 1617-1633 e22. 36. Michealraj, K.A., et al., Metabolic Regulation of the Epigenome Drives Lethal Infantile Ependymoma. Cell, 2020. 181(6): p. 1329-1345 e24. 37. Bongaarts, A., et al., Dysregulation of the MMP/TIMP Proteolytic System in Subependymal Giant Cell Astrocytomas in Patients With Tuberous Sclerosis Complex: Modulation of MMP by MicroRNA-320d In Vitro. J Neuropathol Exp Neurol, 2020. 79(7): p. 777-790. 38. Dodgshun, A.J., et al., Germline-driven replication repair-deficient high-grade gliomas exhibit unique hypomethylation patterns. Acta Neuropathol, 2020. 140(5): p. 765-776. 39. Yu, K., et al., PIK3CA variants selectively initiate brain hyperactivity during gliomagenesis. Nature, 2020. 578(7793): p. 166-171. 40. Monje, M., et al., Roadmap for the Emerging Field of Cancer Neuroscience. Cell, 2020. 181(2): p. 219-222. 41. Caruso, F.P., et al., A map of tumor-host interactions in glioma at single-cell resolution. Gigascience, 2020. 9(10). 42. Voisin, M.R., et al., Brain tumors and COVID-19: the patient and caregiver experience. Neurooncol Adv, 2020. 2(1): p. vdaa104. 43. Mehta, V., et al., Case Fatality Rate of Cancer Patients with COVID-19 in a New York Hospital System. Cancer Discov, 2020. 10(7): p. 935-941. 44. Mohile, N.A., et al., Urgent Considerations for the Neuro-oncologic Treatment of Patients with Gliomas During the COVID-19 Pandemic. Neuro Oncol, 2020. 45. Henriksen, J., et al., Rapid Validation of Telepathology by an Academic Neuropathology Practice During the COVID-19 Pandemic. Arch Pathol Lab Med, 2020. 144(11): p. 1311-1320. 46. Hanna, M.G., et al., Validation of a digital pathology system including remote review during the COVID-19 pandemic. Mod Pathol, 2020. 33(11): p. 2115-2127. 47. Stathonikos, N., et al., Digital pathology in the time of corona. J Clin Pathol, 2020. 73(11): p. 706-712. 48. Williams, B.J., et al., Guidance for Remote Reporting of Digital Pathology Slides During Periods of Exceptional Service Pressure: An Emergency Response from the UK Royal College of Pathologists. J Pathol Inform, 2020. 11: p. 12. 49. Cimadamore, A., et al., Digital pathology and COVID-19 and future crises: pathologists can safely diagnose cases from home using a consumer monitor and a mini PC. J Clin Pathol, 2020. 73(11): p. 695-696. 50. Parker, E.U., O. Chang, and L. Koch, Remote Anatomic Pathology Medical Student Education in Washington State. Am J Clin Pathol, 2020. 154(5): p. 585-591. 51. Mukhopadhyay, S., et al., Leveraging Technology for Remote Learning in the Era of COVID-19 and Social Distancing. Arch Pathol Lab Med, 2020. 144(9): p. 1027-1036. 52. Albeniz, E., et al., Stay connected and up to date. GI meetings and seminars and the COVID-19 pandemic. Tech Innov Gastrointest Endosc, 2020. 53. Bakouny, Z., et al., COVID-19 and Cancer: Current Challenges and Perspectives. Cancer Cell, 2020. 38(5): p. 629-646. 54. Kruger, J. and D. Dunning, Unskilled and unaware of it: how difficulties in recognizing one's own incompetence lead to inflated self-assessments. J Pers Soc Psychol, 1999. 77(6): p. 1121-34. 55. Stichel, D., et al., Accurate calling of KIAA1549-BRAF fusions from DNA of human brain tumours using methylation array-based copy number and gene panel sequencing data. Neuropathol Appl Neurobiol, 2020. 56. Deng, M.Y., et al., Molecularly defined diffuse leptomeningeal glioneuronal tumor (DLGNT) comprises two subgroups with distinct clinical and genetic features. Acta Neuropathol, 2018. 136(2): p. 239-253.
Copyright: © 2021 The author(s). This is an open access article distributed under the terms of the Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited, a link to the Creative Commons license is provided, and any changes are indicated. The Creative Commons Public Domain Dedication waiver (https://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated. |